
PHOTOSYNTHESIS IN HIGHER PLANTS
INTRODUCTION
- All living organisms require energy to carry out their different life activities. This energy directly or indirectly comes from the sun.
- The green plants are unique in the respect that they synthesize food in the presence of sunlight from simple substances like carbon dioxide and water.
- Photosynthesis is the only process of biological importance that can harvest this energy.
- Energy stored as chemical energy in food (carbohydrates, fats and proteins) by photosynthesis is released in living cells during the process of respiration and is utilized to generate heat or converted into energy rich compounds (e.g., ATP and NADP).
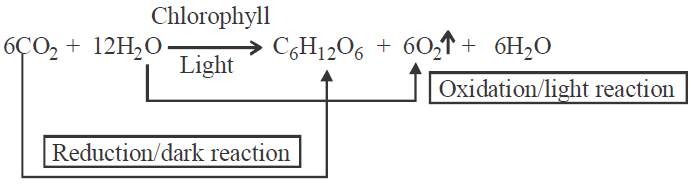
- Photosynthesis is a biochemical process (anabolic and endergonic) in which organic compounds (carbohydrates) are synthesised from the inorganic raw materials (H2O and CO2) in presence of light and pigments. O2 is evolved as a by product.
- 90% of total photosynthesis is carried out by aquatic plants.
- First true and oxygenic photosynthesis started in cyanobacteria (Blue - green algae).
- In the Cuscuta (parasitic) and fungi (achlorophyllous), photosynthesis is absent.
- Euglena is a photosynthetic organism and is the link between animals and plants. Though, they are photosynthetic in the presence of sunlight, when deprived of sunlight they behave like heterotrophs by predating on other smaller organisms.
- Roots of Tinospora and Trapa are assimilatory or photosynthetic.
HISTORICAL BACKGROUND
Before the 17th century, it was considered that plants take their food from the soil.
- Van Helmont (1648) concluded that "all food of the plant is derived from water and not from soil".
- Stephen Hales (1727) reported that plants obtain a part of their nutrition from air and light may also play a role in this process.
- In 1772, Joseph Priestly, carried out some interesting experiments. He took a half filled pot of water and placed a burning candle in its center and covered it with a bell-jar. After sometime, the candle extinguished. Priestly kept a living mouse under the jar and noted that it died immediately. Later he placed a mouse and a potted mint plant under the bell-jar and found that the mouse survived for a long time. He also found that if a mint plant was kept under a bell-jar for a long time and then a burning candle was placed under the jar it continued to burn for a long time. On the basis of these experiments, he concluded that "plants have the capacity to purify foul air".
- Jan Ingen-Housz (1779) concluded by his experiment that purification of air was done by green parts of plant only and that too in the presence of sunlight.
- Jean Senebier (1782) proved that plants absorb CO2 and release O2 in the presence of light. He also showed that the rate of O2 evolution depends upon the rate of CO2 consumption.
- Nicolus de Saussure (1804) showed the importance of water in the process of photosynthesis. He further showed that the amount of CO2 absorbed is equal to the amount of O2 released.
- Julius Robert Mayer (1845) proposed that light has radiant energy and this radiant energy is converted to chemical energy by plants, which serves to maintain the life of the plants and also animals.
- Julius Von Sachs (1862) demonstrated that the first visible product of photosynthesis is starch. He also showed that chlorophyll is confined to the chloroplasts. Thus, he led the foundation of the modern concept of photosynthesis.
- An interesting experiment done by T.W. Engelmann (1843 – 1909) by using a prism showed that it split light into its spectral components and then illuminated a green alga, Cladophora, placed in a suspension of aerobic bacteria. The bacteria were used to detect the sites of O2 evolution.
He observed that the bacteria accumulated mainly in the region of blue and red light of the split spectrum. A first action spectrum of photosynthesis was thus described. It resembles roughly the absorption spectra of chlorophyll a and b.
- In 1930, C.B. van Niel proved the discovery of Nicolas de Saussure wrong. He showed that sulphur bacteria used H2S (in place of water) and CO2 to synthesize carbohydrates as follows :
6CO2 + 12H2S  C6H12O6 + 6H2O + 12S
C6H12O6 + 6H2O + 12S
This led van Niel to the postulation that in green plants, water (H2O) is utilized in place of H2S and O2 is evolved in place of sulphur (S).
6CO2 + 12H2O  C6H12O6 + 6H2O + 6O2.
C6H12O6 + 6H2O + 6O2.
- The discovery of van Niel confirmed by Ruben and Kamen in 1941 using Chlorella, a green alga. He used water (H2O), having heavy isotope of oxygen (O18), oxygen of 18 atomic weight (normal oxygen is of 16 atomic weight) and found that oxygen released in the process of photosynthesis was of O18 type (i.e., heavy oxygen). On the contrary, when CO218 was used, the released oxygen was of normal type, i.e., O216. From this it can be concluded that, oxygen released by green plants comes from splitting of water (oxidation of water).
- Huber, Michel and Deisenhofer (1985) crystallised the photosynthetic reaction center from the purple photosynthetic bacterium, Rhodopseudomonas viridis. They analysed its structure by X-ray diffraction technique. In 1988, they were awarded the Nobel prize in chemistry for this work.
PHOTOSYNTHESIS IN HIGHER PLANTS
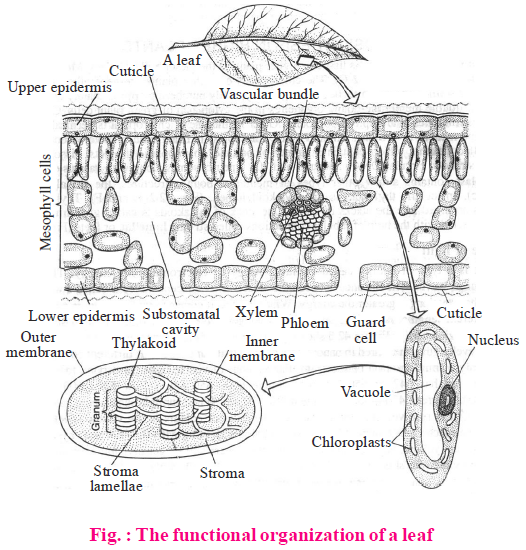
- In higher plants photosynthesis occurs particularly in specialized cells called mesophyll cells of leaves. These cells contain chloroplasts, which are the actual sites for photosynthesis. It fixes CO2 into carbohydrates.
- Chloroplasts are double membrane bound organelles. The space limited by the inner membrane of the chloroplast is called the stroma.
- A number of organised flattened membranous sacs (called the thylakoids) are present in the stroma. Thylakoids are arranged in stacks like piles of coins called grana.
- The thylakoids in the chloroplast contain most of the machinery for the photochemical reaction of photosynthesis. They contain pigments for capturing solar energy.
PHOTOSYNTHETIC PIGMENTS
- Pigments are organic molecules that absorb light of specific wavelengths in the visible region due to the presence of conjugated double bonds in their structures.
- Many pigments are present in photosynthetic cells. PSU (Photosynthetic units) present on thylakoid membranes, are made up of 230-400 molecules of various pigments, called quantasomes, by Park and Biggins.
- Chlorophylls are tetrapyrrolic magnesium (Mg) porphyrin compounds.
- Chlorophyll molecule has a Mg-porphyrin head and an alcoholic phytol tail. Head is hydrophilic and phytol tail is hydrophobic in nature.
- Chlorophyll a – C55H72O5N4Mg (CH3 group at IIIrd C of IInd pyrrole ring)
Chlorophyll b – C55H70O6N4Mg (CHO group at IIIrd C of IInd pyrrole)
Chlorophyll c – C55H32O5N4Mg,
Chlorophyll d – C54H70O6N4Mg,
Carotenes – C40H56
Xanthophylls – C40H56O2.
Chlorophyll d – C54H70O6N4Mg,
Carotenes – C40H56
Xanthophylls – C40H56O2.
- Chl-a and carotenes are universal pigments, which are found in all O2 liberating cells.
- Chlorophylls are soluble only in organic solvents like ketones, ethers etc.
- Stroma lamellae/stroma thylakoids lack PS II and enzyme NADP reductase.
- By paper chromatography/chromatogram, produces different pigment colours which appear as:
- Chlorophyll a - blue-green.
- Chlorophyll b - yellow green.
- Xanthophyll - yellow
- Carotenoids - yellow to yellow - orange
- Carotenoids absorb light energy and transfer it to Chl a and thus, act as accessory pigments. They protect the chlorophyll molecules from photo-oxidation by picking up nascent oxygen and converting it into harmless molecular stage. Carotenoids can be classified into two groups namely carotenes and xanthophylls.
- Carotenes : They are orange red in colour and have general formula C40H56. They are isolated from carrot. They are found in all groups of plants i.e., from algae to angiosperms. Some of the common carotenes are α, β, γ and δ carotene; phytotene, lycopene, neurosporene etc. Lycopene is a red pigment found in ripe tomatoes and red pepper fruits. The β-carotene on hydrolysis gives vitamin A, hence the carotenes are also called provitamin A. β-carotene is black yellow pigment of carrot roots.
- Xanthophylls : They are yellow coloured carotenoid also called as xanthols or carotenols. They contain oxygen also along with carbon and hydrogen and have general formula C40H56O2.
Lutein (C40H56O2) is a widely distributed xanthophyll which is responsible for yellow colour in autumn foliage. Fucoxanthin (C40H56O6) is an another important xanthophyll present in Phaeophyceae (brown algae).
- Phycobilins : These pigments are mainly found in blue-green algae (cyanobacteria) and red algae. These pigments have open tetrapyrrolic structure and do not bear magnesium and phytol chain.
- Blue-green algae have higher quantity of phycocyanin and red algae have more phycoerythrin. Phycocyanin and phycoerythrin together form phycobilins. These water soluble pigments are thought to be associated with small granules attached with lamellae. Like carotenoids, phycobilins are accessory pigments i.e., they absorb light and transfer it to chlorophyll a.
ABSORPTION AND ACTION SPECTRA
The curve representing the light absorbed at each wavelength by pigment is called absorption spectrum. Curve showing rate of photosynthesis at different wavelengths of light is called action spectrum.
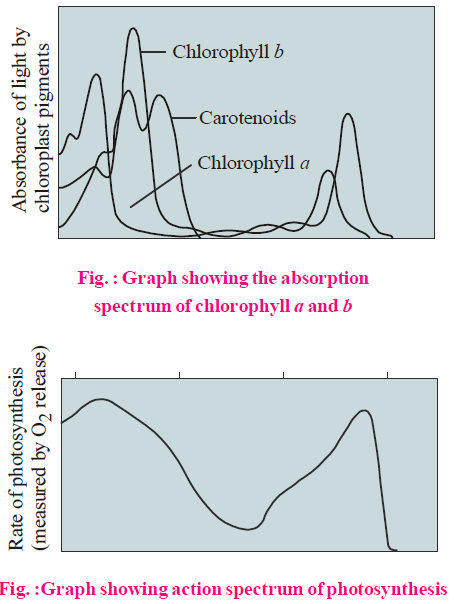
Absorption spectrum is studied with the help of spectrophotometer. The absorption spectrum of chlorophyll a and chlorophyll b indicate that these pigments mainly absorb blue and red lights, (430 nm and 662 nm for chlorophyll a, 455 nm and 644 nm for chlorophyll b). Action spectrum shows that maximum photosynthesis takes place in blue and red regions of the spectrum. The first action spectrum of photosynthesis was studied by T.W. Engelmann (1882) using green alga Spirogyra and oxygen seeking bacteria.
MECHANISM OF PHOTOSYNTHESIS
- Photosynthesis is an oxidation reduction process in which water is oxidised to release O2 and CO2 is reduced to form starch and sugars.
- Scientists have shown that photosynthesis is completed in two phases –
- Light phase or photochemical reactions or light dependent reactions or Hill's reactions : During this stage, energy from sunlight is absorbed and converted to chemical energy which is stored in ATP and NADPH + H+.
- Dark phase or chemical dark reactions or light independent reactions or Blackmann reaction or biosynthetic phase : During this stage, carbohydrates are synthesized from carbon dioxide using the energy stored in the ATP and NADPH formed in the light dependent reactions.

LIGHT PHASE OR PHOTOCHEMICAL REACTIONS OR LIGHT DEPENDENT REACTIONS OR HILL'S REACTIONS
Light reaction occurs in the grana fraction of the chloroplast and in this reaction are included those activities, which are dependent on light. Assimilatory powers (ATP and NADPH2) are mainly produced in this light reaction.
Robin Hill (1939), first of all showed that if chloroplasts extracted from leaves of Stellaria media and Lamium album are suspended in a test tube containing suitable electron acceptors, e.g., potassium ferroxalate (some plants require only this chemical) and potassium ferricyanide, oxygen is released due to photochemical splitting of water. Under these conditions, no CO2 was consumed and no carbohydrate was produced, but light-driven reduction of the electron acceptors was accompanied by O2 evolution.
The splitting of water during photosynthesis is called photolysis. This reaction is known as Hill reaction.
Hill as reaction proves that
- In photosynthesis, oxygen is released from water.
- Electrons for the reduction of CO2 are obtained from water [i.e., a reduced substance (hydrogen donor) is produced which later reduces CO2].
2, 6 – dichlorophenolindophenol (DCPIP) is the dye used by Hill for his famous Hill's reaction.
It is a blue chemical compound used as a redox dye. This dye changes colour (if reduced, colourless) when exposed to light in a photosynthetic system.
According to Arnon (1961), in this process, light energy is converted to chemical energy. This energy is stored in ATP (this process of ATP formation in chloroplasts is known as photophosphorylation) and from electron acceptor NADP+, a substance found in all living beings NADPH is formed as hydrogen donor. Formation of hydrogen donor NADPH from electron acceptor NADP+ is known as photoreduction or production of reducing power NADPH.
QUANTUM YIELD
- Rate or yield of photosynthesis is measured in terms of quantum yield or O2 evolution, which may be defined as, “Number of O2 molecules evolved per quantum of light absorbed in photosynthesis.”
- Quantum requirement in photosynthesis = 8, i.e., 8 quanta of light are required to evolve one molecule of O2.
- Hence, quantum yield = 1 / 8 = 0.125 (i.e., a fraction of 1) as 12%.
EMERSON EFFECT AND RED DROP
R. Emerson and C.M. Lewis (1943) observed that the quantum yield of photosynthesis decreases towards the far red end of the spectrum (680nm or longer). Since this decrease in quantum yield is observed at the far region or beyond red region of spectrum it is called red drop.
Emerson et al. (1957) further observed that photosynthetic efficiency of light of 680nm or longer is increased if light of shorter wavelengths (less than 680nm) is supplied simultaneously. When both short and long wavelengths were supplied together the quantum-yield of photosynthesis was greater than the total effect when both the wavelengths were supplied separately. This increase in photosynthetic efficiency (or quantum yield) is known as Emerson effect or Emerson enhancement effect.

PIGMENT SYSTEMS
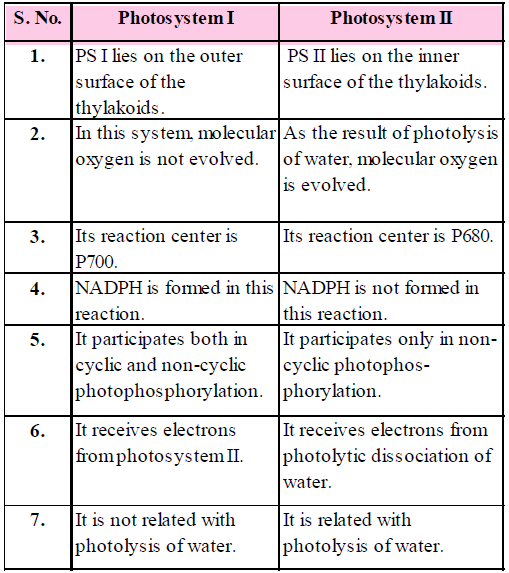
- The pigments are organised into two discrete photochemical light harvesting complexes (LHC) within Photosystem I (PS I) and Photosystem II (PS II).
- These pigments help to make photosynthesis more efficient by absorbing different wavelengths of light. The single chlorophyll a molecule forms the reaction centre. The reaction centre is different in both the photosystems. In PS I, the reaction centre chlorophyll a has an absorption peak at 700 nm, hence is called P700, while in PS II it has absorption maxima at 680 nm, and is called P680.
- Pigment system I or Photosystem I (PS I) : The important pigments of this system are chlorophyll a 670, chlorophyll a 683, chlorophyll a 695, P700. Some physiologists also include carotenes and chlorophyll b in pigment system I. Thus, this system absorbs both wavelengths that are shorter and longer than 680nm.
- Pigment system II or Photosystem II (PS II) : The main pigments of this system are chlorophyll a 673, P680, chlorophyll b and phycobilins. This pigment system absorbs wavelengths shorter than 680nm only.
Table : Comparison between Photosystem I and Photosystem II
- Antenna or accessory pigments receive radiant energy and transfer it among themselves. This transfer of energy is known as resonance transfer. Then antenna gets molecules excited and transfer their energy to chlorophyll 'a' molecules of reaction centre. It is known as inductive resonance. Finally chl. 'a' molecules convert the light energy into electrical energy by bringing about electric charge separation.
CYCLIC ELECTRON TRANSPORT SYSTEM
CYCLIC PHOTOPHOSPHORYLATION
- In cyclic photophosphorylation, only PS-I (LHC-I) works.
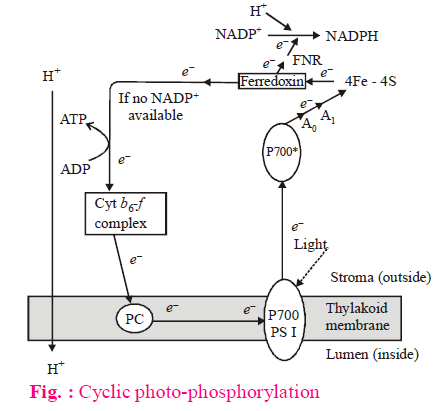
- Cyclic ETS or PS-I is activated by wavelength of light greater than 680 nm.
- It occurs at grana thylakoids and stroma thylakoids.
- During cyclic ETS, the electron ejected from reaction centre of PS-I, returns back to its reaction centre.
- In cyclic ETS, no oxygen evolution occurs because photolysis of water is absent.
- Phosphorylation takes place at two places, thus two ATP are generated in each cyclic ETS.
- NADPH2 (reducing power) is not formed in cyclic process.
- Plastocyanin (PC) is a Cu-containing blue protein in cyclic ETS.
- According to modern researches, first e– acceptor is FRS (Ferredoxin Reducing Substance), which is a Fe-S containing protein. Earlier fd (Ferredoxin) was considered as the first e– acceptor.)
Z-SCHEME/NON-CYCLIC ELECTRON TRANSPORT SYSTEM/NON CYCLIC PHOTOPHOSPHORYLATION
- Both PS-I and PS-II is involved in non-cyclic ETS.
- It occurs at grana thylakoids only.
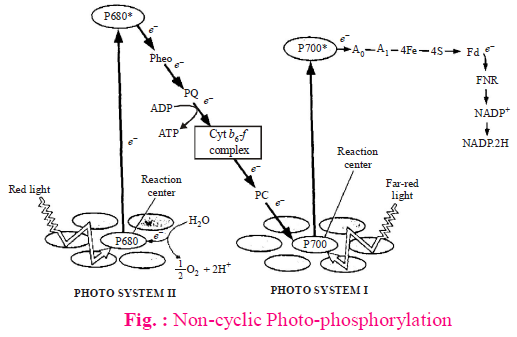
- The e– ejected from PS-II never goes back to chla-680 (reaction centre) and finally gained by NADP. During this process oxygen is evolved due to photolysis of water and formation of NADPH2 and ATP occurs.
- Each turn of non-cyclic ETS produces 1 ATP and 2NADPH2 (4 molecules of H2O are photolysed and
1 molecule of O2 is released). - Primary e– acceptor in noncyclic reaction is PQ or plastoquinone. Recently pheophytin (structure like chla without Mg) is considered as the first e– acceptor in Z-scheme.
- Plastocyanin (Cu-containing pigment) is the link between PS-I and PS-II in non-cyclic ETS.
- Final e– acceptor in Z-scheme is NADP+ (Hills reagent).
- During non-cyclic ETS energy flow takes place from PS II to PS I.
- Non-cyclic photophosphorylation or Z-scheme is inhibited by 3-(4-chlorophenyl)-1, 1-dimethylurea (CMU) and 3-(3,4-dichlorophenyl)-1,1 dimethylurea (DCMU) is an herbicide which kills the weeds by inhibiting CO2 fixation as it is a strong inhibitor of PS-II.
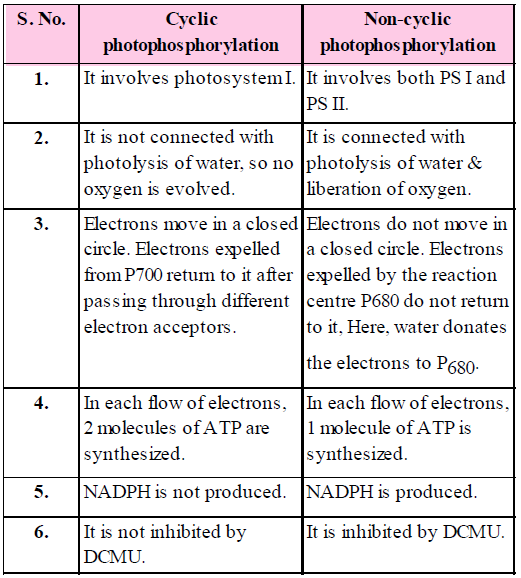
CHEMIOSMOTIC HYPOTHESIS
- Chemiosmotic hypothesis has been put forward by Mitchell (1961) to explain the mechanism of ATP synthesis.
- When electrons are transported through the electron transport system (ETS), the protons accumulate inside the thylakoid membranes.
- Lumen of thylakoid becomes enriched with H+ ion due to photolytic splitting of water.
- Primary acceptor of electron is located on the outer side of the thylakoid membrane. It transfers its electron to an H-carrier. The carrier removes a proton from the matrix while transporting electrons to the inner side of the membrane. The proton is released into the lumen while the electron passes to next carrier.
- NADP reductase is situated on the outer side of the thylakoid membrane. It obtains electrons from PSI and protons from matrix to reduce NADP+ to NADP + H+ state.
- The proton gradient is broken down due to the movement of protons through transmembrane channels, F0 of ATPase. ATPase enzyme has two parts – F0 and F1 particles.
- The rest of the membrane is impermeable to H+.
- F0 provides facilitated diffusion to H+ or protons. As protons move to the outer side of ATP, they bring about conformational changes in F1 particle of ATPase or coupling factor. The transient F1 particle of ATPase enzyme form ATP from ADP and inorganic phosphate.
- One molecule of ATP is formed when 2H+ ions pass through ATPase.
DARK PHASE OR CHEMICAL DARK REACTIONS OR LIGHT INDEPENDENT REACTIONS OR BLACKMANN REACTION OR BIO-SYNTHETIC PHASE
- The pathway by which all photosynthetic eukaryotic organisms ultimately incorporate CO2 into carbohydrate is known as carbon fixation or photosynthetic carbon reduction (PCR) cycle or dark reaction.
- The dark reaction is sensitive to temperature changes, but are independent of light hence it is called dark reaction, however, it depends upon the products of light reaction of photosynthesis, i.e., NADP. 2H+ and ATP.
- Carbon dioxide fixation takes place in the stroma of chloroplasts because it has enzymes essential for fixation of CO2 and synthesis of sugar.
CALVIN CYCLE / C3-CYCLE
- Calvin and Benson discovered the path of carbon in this process. First stable compound of Calvin cycle is 3C-PGA (phosphoglyceric acid), thus Calvin cycle is called as C3-cycle.
- It was studied by Calvin on Chlorella and Scenedesmus These are microscopic unicellular algae and can be easily maintained in the laboratory. During his experiment, he used chromatography and radioactive tracer (C14) techniques for detecting reactions of C3-cycle.
- RuBisCO (Ribulose bis-phosphate carboxylase oxygenase) is the main enzyme in C3-cycle, which is present in the stroma and it makes about 16% protein of chloroplast. RuBisCO is the most abundant enzyme on earth.
- CO2-acceptor in C3 cycle is Ribulose 1,5- diphosphate (RuBp). This carboxylation reaction is catalysed by RuBisCO.
- Atriplex hastata and A. patula are temperate species, which are C3-plants.
- 6 turns of Calvin cycle are required for the formation of one glucose.
- In this cycle, for formation of one mole of hexose sugar (glucose), 18 ATP and 12 NADPH2 are used.
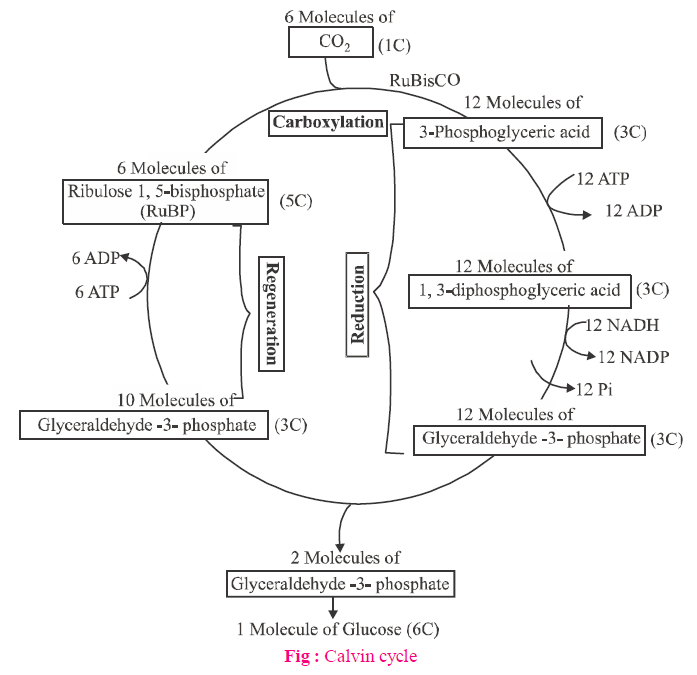
Calvin cycle is divided into three distinct phases : carboxylation, glycolytic reversal and regeneration of RuBP.
(1) Carboxylation
(2) Glycolytic Reversal
12 mol. of 1, 3-BiPGA 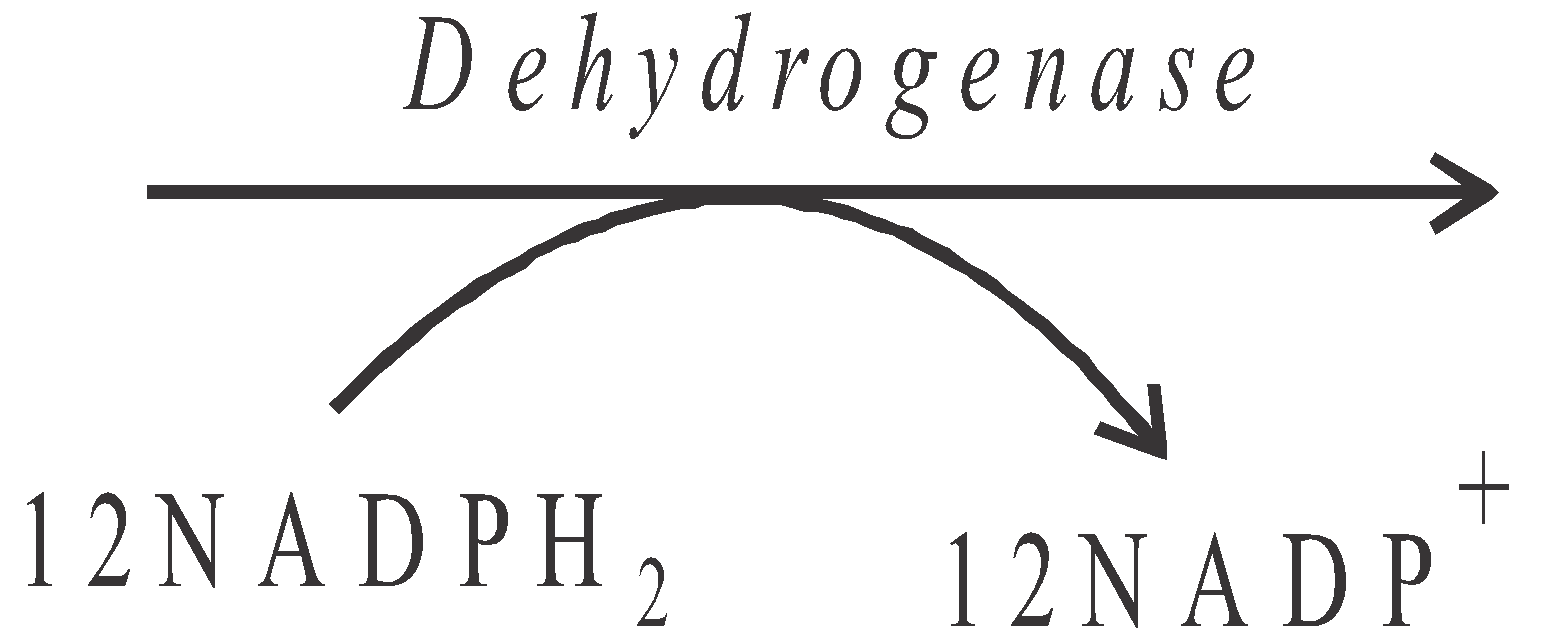 12 mol. of 3–phosphoglyceraldehyde (3-PGAL) (Triose phosphate) + 12H3PO4
12 mol. of 3–phosphoglyceraldehyde (3-PGAL) (Triose phosphate) + 12H3PO4
Out of these 12 molecules, 2 molecules go to synthesize sugar, starch and other carbohydrates and the remaining 10 molecules are recycled to regenerate 6 molecules of Ribulose-5-phosphate after a series of complex reactions.
Out of 2 molecules of PGAL, one molecule is converted to its isomer 3-hydroxy acetone phosphate.
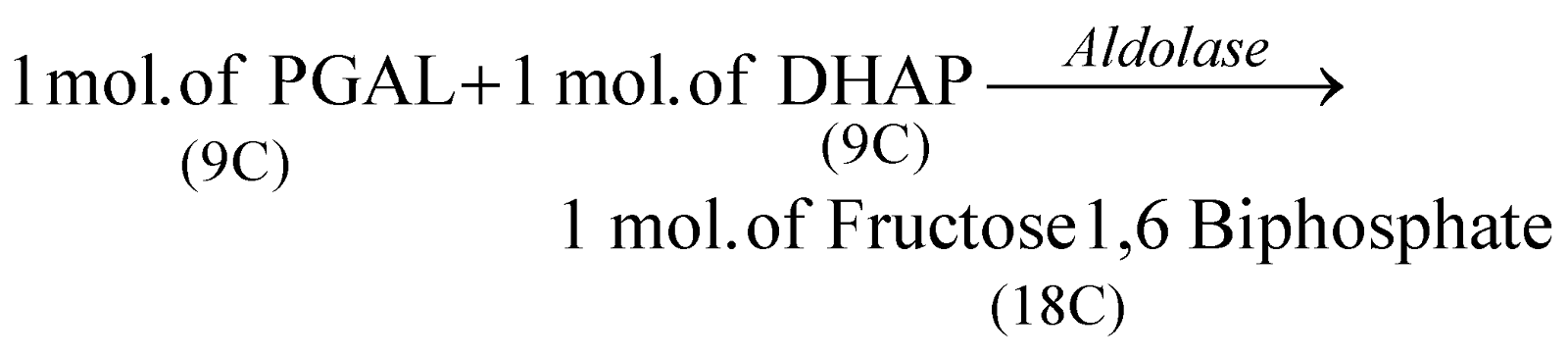
[1 mol. fructose  [C6H12O6] Glucose
[C6H12O6] Glucose  Sucrose/Starch]
Sucrose/Starch]
(3) Regeneration of Ribulose 1,5 biphosphate
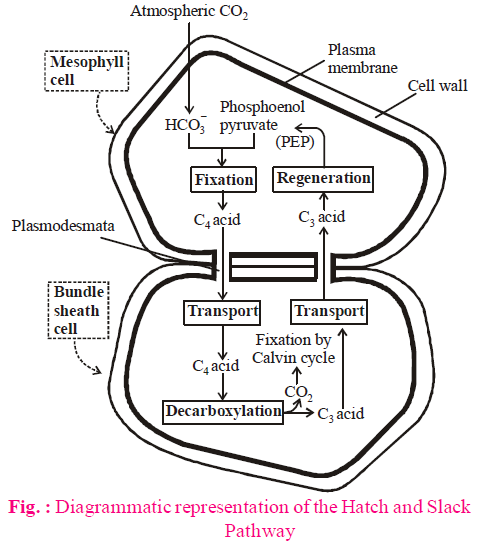
C4 CYCLE (HATCH & SLACK PATHWAY)
- Kortschak and Hartt first observed that 4C-compound OAA (Oxalo Acetic Acid) is formed during dark reaction in sugarcane leaves.
- Hatch and Slack (1967) studied in detail and proposed a pathway for dark reactions in sugarcane and maize leaves.
- First stable product of this reaction is OAA, which is a 4C compound, a dicarboxylic substance, thus Hatch and Slack pathway is called as C4 cycle or dicarboxylic acid cycle (DCA).
- C4-cycle occurs in 1500 species of 19 families of angiosperms, but most of the plants are monocots, which belongs to Gramineae and Cyperaceae (sugarcane, maize, sorghum, oat, chloris, sedges, bajra, Panicum, Alloteropsis etc.)
- Dicots with C4-cycle are Euphorbia spp., Amaranthus, Chenopodium, Boerhavia, Atriplex rosea, Portulaca, Tribulus.
- Kranz (Wreath) Anatomy - Present in leaves of C4 plants.
- There are various features of Kranz anatomy which are as given below :
- Green bundle sheath cells (BS cells) are present around the vascular bundles.
- Dimorphic chloroplasts are present in leaf cells. Chloroplast of bundle sheath cells or Kranz cells are larger and lack grana. Mesophyll chloroplasts are small and with grana.
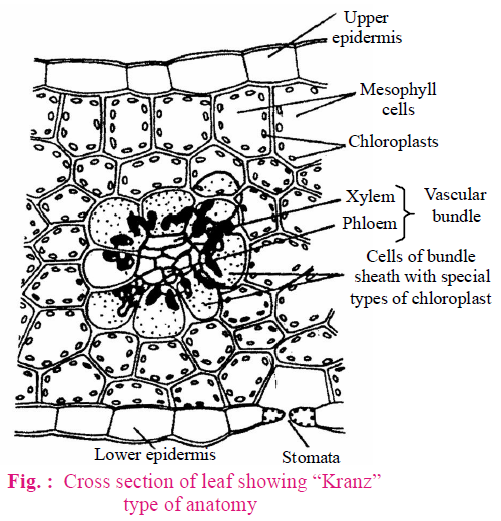
- In the C4-plant, C3-cycle occurs in the bundle sheath cells, while C4-cycle occurs in the mesophyll cells.
- Operation of Hatch and Slack pathway requires cooperation of both photosynthetic cells i.e. mesophyll cells and bundle sheath cells.
- Photosynthetically, C4 plants are more efficient as there is no Warburg effect or photorespiration in C4-plants, because at the site of RuBisCo (BS cells), less O2 is present (mesophyll cells pump more CO2 for C3 cycle).
- C4-plants are found in tropical habitats and have adapted themselves in high temperature, low water availability and intense light. So, they are more developed and adapted than C3 plants. They do not cause any loss to photorespiration.
- If concentration of O2 increases artificially, then photorespiration may be started in C4 plants.
- First, carboxylation in C4-cycle occurs by PEPCase in mesophyll chloroplast, while second carboxylation or final CO2 fixation by C3 cycle occurs in bundle sheath cells. PEPCase is an enzyme in the carboxylase that is responsible for the formation of the 4-C compound oxaloacetate by catalyzing the binding of one molecule of CO2 to the 3C compound phosphoenolpyruvate.
- CO2 acceptor in C4 mesophyll cell is phosphoenolpyruvate (PEP), a 3C-compound, while RuBp in bundle sheath cells.
- 12 NADPH2 (= 36 ATP) + 30 ATP = 66 ATP are needed for production of 1 hexose (glucose) in C4-plants.
- Pyruvate phosphate dikinase (PPDK) (that converts ATP into AMP) is a temperature sensitive enzyme of C4 and CAM plants due to which C4 plants photosynthesize better at high temperature.
Special features of C4 plants :
- C4 plants are more efficient plants at a given CO2 concentration.
- Level of atmospheric CO2 present is generally not a limiting factor for C4 plants.
- C4 plants possess low CO2 compensation points. (8-10 ppm)
- The productivity (fertility) does not increase in C4 plants, when CO2 concentration increases because-
- Mesophyll cells provide more CO2 for the Calvin cycle.
- The concentration of CO2 around the site of RuBisCO is higher in C4 plants, thus there is little or no chance of photorespiration.
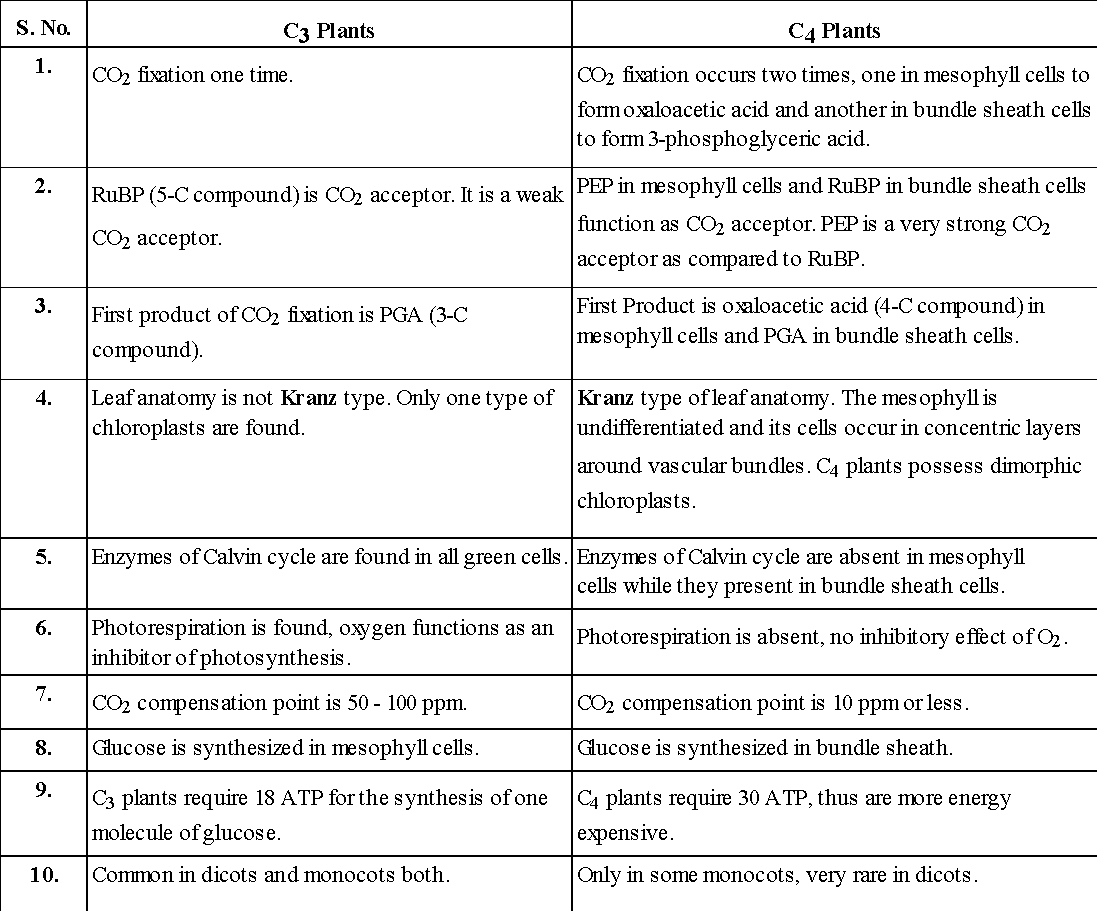
Table : Comparison between C3 plants and C4 plants
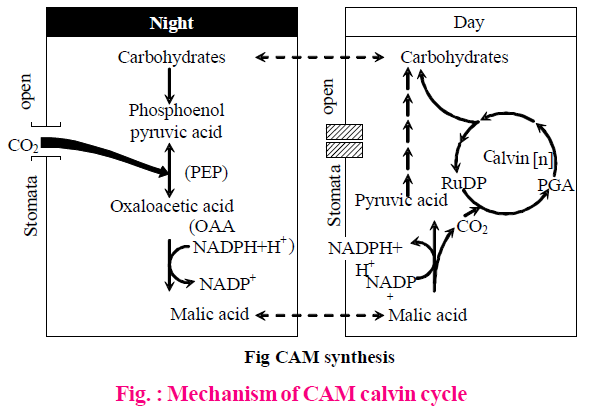
CAM (CRASSULACEAN ACID METABOLISM) PLANTS
- This dark CO2 fixation pathway was proposed by Ting (1971).
- It operates in succulent or fleshy plants e.g., Kalanchoe, Crassula, Sedum, Opuntia, Aloe, Agave, Euphorbia sps., Bryophyllums, Pineapple, Welwitschia (gymnosperm) etc.
- Primary acceptor of CO2 is PEP (Phosphoenol pyruvate) and oxaloacetic acid is the first product of carboxylation reaction.
- In CAM plants, stomata are of scotoactive (active during night) type, so initial CO2 fixation is found in night but light reactions operate at day time. Final CO2 fixation (C3 cycle) occurs in day time. PEPCase induces carboxylation reaction at night.
- PEP carboxylase and RuBisCO are present in mesophyll cells. (No Kranz-anatomy)
- In CAM plants, 30 ATP and 12 NADPH2 are required for synthesis of 1 glucose molecule.
- Formation of malic acid during dark phase is called acidification or phase-I. Release of CO2 for actual photosynthesis during day is called deacidification or phase-II.
- Malic acid is stored in the vacuoles during the night which is decarboxylated to release CO2 during the day.
CO2 COMPENSATION POINT
- In photosynthesis, CO2 is utilized in the presence of light to release O2 whereas in respiration, O2 is taken and CO2 is released. If light factor is saturating, there will be certain CO2 concentration at which the rate of photosynthesis is just equal to the rate of respiration, or photosynthesis just compensates respiration or apparent photosynthesis is nil. It is called CO2 compensation point.
- CO2 compensation point is very low in C4 plants, i.e, 0 to 5 ppm whereas high CO2 compensation point is found in C3 plants, i.e., 25 to 100 ppm.
PHOTORESPIRATION / C2 CYCLE / GLYCOLATE PATHWAY
Krotkov indicated that more CO2 evolves during day time in C3 plants.
- Decker and Tio discovered photorespiration and clarified that C2-cycle or glycolate pathway operates during day time in C3-plants and RuBisCO acts as oxygenase at higher concentration of O2 and low CO2 concentration in the C3 - green cells.
- The light dependent uptake of O2 and release of CO2 in C3 photosynthetic cell is called photorespiration.
- Photorespiration is not linked with ATP generation (in place ATP are consumed) as ordinary dark respiration, thus it is harmful or wasteful process linked with C3 cycle.
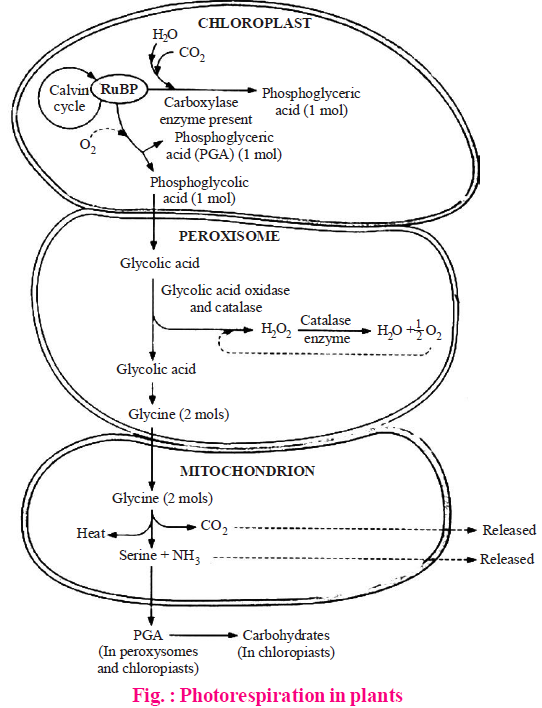
- It is assumed that in C3 plants, if photorespiration does not occur, it will increase O2 conc. which oxidises (photooxidation) the different protoplasmic parts of photosynthetic cell at high light intensity.
- Photorespiration is enhanced by bright light, high temperature, high oxygen and low CO2 concentration.
FACTORS AFFECTING PHOTOSYNTHESIS
There are several external and internal factors which affects the rate of photosynthesis.
EXTERNAL FACTORS
- Light :
Light Quality or wavelength : Maximum photosynthesis takes place in red light than blue light. But the rate of photosynthesis is highest in white light while minimum is in green light.
Light Intensity : Rate of photosynthesis is greater in intense light than in diffused light. But at higher light intensity, photooxidation (solarization) occurs and photosynthetic apparatus may get destroyed.
P/R (Photosynthesis: Respiration) Ratio at mid day is 10 : 1; but can reach upto 20 : 1. At evening and morning, the rate of photosynthesis equals to respiration, this situation is called as light compensation point.
Plants which are adapted to grow in high intensity of light are called heliophytes and plants which are adapted to grow in shade are called sciophytes.
- Temperature : Optimum temperature for photosynthesis is 20-35°C. At high temperature, the rate of photosynthesis decreases due to denaturation of enzymes.
- CO2 : An increase in CO2 concentration upto 1% (0.03% to 1%) causes rate of photosynthesis to increase. Higher CO2 concentration is toxic to plants and also closes stomata.
- O2 : High O2 concentration reduces photosynthesis due to photorespiration.
Warburg effect : Inhibitory effect of high concentration of O2 on photosynthesis is called as Warburg effect (It is due to photorespiration).
- Water : Less availability of water reduces the rate of photosynthesis (stomata get closed).
- Inhibitors : DCMU (diuron/dichlorophenyl dimethyl Urea), CMU (Monuron), PAN, atrazine, simazine, bromacil inhibit photosynthesis by blocking PS-II. They stop electron flow between P-680 and PQ.
In cyclic ETS, diquat, paraquat (Viologen dyes) inhibit electron flow between P-700 and Fd. All these chemicals are used as herbicides, which mostly block electron transport system (ETS).
- Minerals : Magnesium and nitrogen are essential for the structure of chlorophyll and enzymes. Thus, reduction in N2 and Mg supply to plants affects the rate of photosynthesis.
RuBisCO alone accounts for more than half of the total leaf nitrogen. Generally, all essential elements affect the rate of photosynthesis.
INTERNAL FACTORS
- Chlorophyll : The amount of CO2 in grams absorbed by 1 gm. of chlorophyll in 1 hour is called as photosynthetic number or assimilatory number.
- Product of photosynthesis : Rate of photosynthesis decreases, when sugar accumulates in mesophyll cells.
- Leaf : Various leaf factors like leaf age and leaf orientation affect the rate of photosynthesis. In young & mature leaves, photosynthesis is more than old (senescent) leaves.
LAW OF MINIMUM (LIEBIG)
According to it, when a process is governed by a number of separate factors, then the rate of process is controlled by that factor present in minimum amount.
LAW OF LIMITING FACTORS (BLACKMAN)
It is the modification of Law of minimum by Liebig. "When a process is conditioned to its rapidity by a number of factors, then rate of the process is limited by the pace of the slowest factor (CO2, light, chlorophyll, water, temp).
BACTERIAL PHOTOSYNTHESIS
- Certain bacteria are capable of photosynthesis. E.g., Chlorobium (Green Sulphur), Chromatium (Purple Sulphur), Rhodospirillum, Rhodopseudomonas (Purple non sulphur).
- Cyclic-photophosphorylation is an important method in bacterial photosynthesis because PS-II is absent here, therefore, Z-scheme is absent in bacteria.
- Absorption of infrared spectrum takes place during bacterial photosynthesis thus, no red drop occurs in bacterial photosynthesis.
- Pigment system of bacteria is denoted by P-890 or 870.
- Only one ATP is produced in each turn of cyclic photophosphorylation in bacteria.
- Chemosynthesis
The process of carbohydrate formation in which organisms use chemical reactions to obtain energy from inorganic compounds is called chemosynthesis. Such chemoauto-trophic bacteria do not require light and synthesize all organic cell requirements from CO2 and H2O and salts at the expense of oxidation of inorganic substances like (H2, NO3–, SO42– or carbonate). Some examples of chemosynthesis are Nitrosomonas, Nitrobacter, Beggiatoa, Thiothrix, Ferrobacillus, Bacillus pantotrophus, Carboxy Dominos etc.
SIGNIFICANCE OF PHOTOSYNTHESIS
- Photosynthesis is a vital process for life on planet earth as it is the only process that links the physical and biological world by conversion of solar energy into organic matter, which makes up the bulk of the dry matter of any organism.
- Presence of O2 in the atmosphere is also an outcome of photosynthesis. This oxygen is helpful to living organisms in two ways:
- Oxidative breakdown of organic food matter (respiration).
- Making ozone (O3) in outer layer of atmosphere which helps in stopping the highly destructive UV rays.
Post a Comment