
ANATOMY OF FLOWERING PLANTS
INTRODUCTION
- Plant anatomy is the branch of botany which deals with study of internal structures and organization of plants.
- Nehemiah Grew is known as the father of plant anatomy.
- K.A. Chaudhary is known as the father of Indian plant anatomy.
TISSUE
- A group of cells which is similar or dissimilar in shape, having a common origin and usually performing a common function is called tissue.
- The term tissue was coined by Nehemiah Grew.
- The tissues are divided into two groups by Carl Nageli on the basis of stage of development– meristematic and permanent.
MERISTEMATIC TISSUE
Meristematic tissues is a simple plant tissue in which growth in plants is largely restricted to specialised regions of active cell division. A meristem is a localised region in which actual cell division occurs.
CHARACTERISTICS OF MERISTEMATIC TISSUES
- Meristematic tissues or meristem is an undifferentiated tissue.
- Cell cycle of meristem is in continuous state of division. It means they have the capacity to divide. So, meristematic tissue is composed of immature cells.
- Meristematic cells have only primary cell wall which is thin and flexible (elastic) and made up of cellulose. Secondary cell wall is absent.
- Cells of meristem are small and isodiametric and they have dense cytoplasm.
- Normally, vacuoles are absent in meristematic cells but if present then small.
- They have prominent and large nucleus.
- Meristematic cells are metabolically highly active so they lack reserve food.
- Plastids are absent in meristems. If they are present, then only in the proplastid stage.
- They do not have intercellular spaces. Cells are closely fitted packed together. So, it is a compact tissue.
CLASSIFICATION OF MERISTEMATIC TISSUE
(A) BASED ON ORIGIN AND DEVELOPMENT
On the basis of origin and development, meristems can be divided into 3 types – promeristem, primary and secondary meristem.
Promeristem/Embryonic Meristem
- This meristem develops in beginning during embryonic stage.
- Promeristems are usually found in root apex or shoot apex.
- They develop primary meristem.
Primary Meristem
- Meristematic cells developed from promeristem are known as primary meristem.
- These cells are always in division phase and form primary permanent tissue.
Secondary Meristem
- These are the meristems developed from primary permanent tissues. They are not present in the embryonic stage of the plant.
- Some of the cells of primary permanent tissues become meristematic and constitute secondary meristem.
- By the activity of secondary meristems, secondary growth takes place.
(B) BASED ON LOCATION (POSITION) IN PLANT BODY
On the basis of position, meristematic tissues are divided into three types : apical, intercalary and lateral meristem.
(1) Apical Meristem
- The meristems which occur at the tips of roots and shoots and produce primary permanent tissues.
- They are responsible for increase in the length of plant organs and primary growth. E.g., Root apex, shoot apex.
- Apical meristem has two regions at embryonic stage : promeristem or primordial meristem which develops in embryonic stage and eumeristem, which is formed from the division of primordial meristem.

According to Haberlandt, eumeristem is divided into three regions (on the basis of function) :
- Protoderm or Dermatogen : It is the outermost layer of eumeristem. By the activity of protoderm, epidermal tissue system is formed. It includes epidermis, root hair, stem hair etc.
- Procambium or Plerome : These cells are long and give rise to the vascular tissue system. It includes xylem and phloem.
- Ground meristem or Periblem : The cells of this region are large, thin walled and isodiametric. Ground tissue system is formed by the activity of these cells. It includes hypodermis, cortex, endodermis, pericycle, pith-rays and pith.

(2) Intercalary Meristem
- This meristem occurs between mature tissues and is the separated region of apical meristem.
- It is found in between the plant organs.
- By the activity of this meristem, length of the plant organ increases.
- They are present in some stem of plants. They may be present either at the base of internode (e.g., grasses, bamboo and Equisetum etc.), or at the base of node, (e.g., Mint). They are also present at the base of leaves, e.g., Pinus. By the activity of this meristem, length of leaves increases.
- These meristems are short lived. On loosing their power of division, they merge with permanent tissue.
(3) Lateral Meristem
- Lateral meristem occurs in lateral side of plant organ or parallel to the longitudinal axis (tangential plane) of plant organ.
- Activity of lateral meristem increases the width of plant organ so it is responsible for secondary growth.
- Lateral meristems are both primary and secondary in origin (mostly secondary in origin). There are two examples of primary lateral meristem :
- Marginal meristem : It occurs at the margin of leaf. Its activity increases the width of leaf, so total growth of leaf is called intercalary marginal growth.
- Intra-fascicular cambium or fascicular cambium : This cambium occurs inside the vascular bundles. Except intrafascicular cambium, all cambia are secondary in origin.
- Lateral meristems are cylindrical.

(C) BASED ON PLANE OF DIVISION
- On the basis of plane of division, meristem can be classified as rib, plate and mass meristem.
- Rib-meristem/File meristem are meristems in which anticlinal division occurs in one plane. For example, tunica is a type of rib-meristem.
Formation of some cells of the cortex and pith takes place by this meristem.
- The meristem which divides anticlinally into two planes at right angles to each other are called plate meristem. By this division, a plate like structure is formed. Formation of leaf blade takes place by the activity of this meristem.
- Mass-meristem divides in all possible planes and resulting in an increase in the volume of plant body. E.g., the formation of embryo and endosperm takes place by this kind of meristem.
Composition of Apical Meristem in Different Plants
- Apical meristem is absent in lower algae and fungi. All the cells of these plants are divisible, so they do not show apical growth. Thus, such type of growth in these plants is called diffused growth. Diffused growth also occur in animals.
- Apical meristem in higher algae (e.g., Fucus, Dictyota & Sargassum), bryophytes and pteridophytes (e.g., Selaginella) consists of a single cell. This cell is known as apical cell. This apical cell is pyramid in shape and divide into two lateral planes.
- Apical meristem in ferns, gymnosperms and angiosperms consists of many cells.
- Several theories have been available in relation to structure and growth habit of apical meristem. Types of theories are – apical cell theory, histogen theory, tunica corpus theory, mantle core theory, Newman’s theory & Korper-Kappa theory.
(1) Apical cell theory
- This theory was proposed by Karl Nageli and Hofmeister and supported by Wolff.
- According to Nageli and Hofmeister, the apical meristem is composed of single apical cell. This view is only applicable on Bryophytes and some Pteridophytes and some higher algae (Fucus, Dictyota & Sargassum).
(2) Histogen Theory
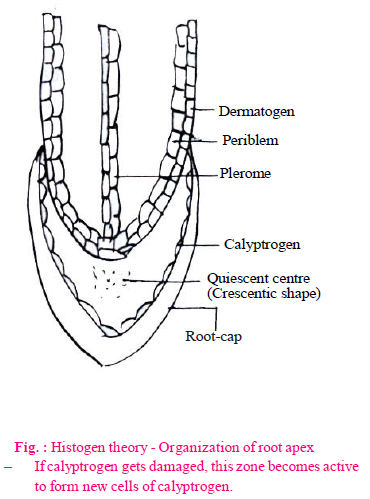
- It was proposed by Hanstein (1870).
- According to him, the root and shoot apices are distinguished into three meristematic regions or three layers of histogen cells. These are as follows–

- Dermatogen : This is the outermost single layer of cells. These cells form uniseriate epidermis by anticlinal divisions.
- Periblem : This region is situated just below the dermatogen. It forms cortex (hypodermis, general cortex and endodermis).
- Plerome : This is the innermost region. Stele formation takes place by division of these cells. It means formation of pericycle, vascular bundles, pith rays or medullary rays and pith takes place.
- This theory is only true for root apex. It is not applicable for shoot apex of higher plants because in most of gymnosperms and angiosperms, shoot apex does not have distinct differentiation of three layers.
- Except above described three histogens, a fourth type of histogen is present in monocotyledons. This is known as calyptrogen. Root cap is produced by calyptrogen in monocots. Root cap is produced by dermatogen in dicotyledons.
Exception : There is only one histogen present in Ranunculus. Two histogens occur in Casuarina.
Due to the presence of root cap, position of root apex is sub terminal, so maximum growth in root takes place behind the apex.
NOTES
- In hydrophytes, root cap is absent.
- Generally root cap is single layered but in Pandanus (screw pine), root cap is multilayered.
- Root cap contain more amount of golgi body which secrete mucilage which make the root slimy.
- A group of inactive cells present between the dermatogen and calyptrogen is called quiescent centre. These cells contain less amount of DNA and synthesis of protein is also less.
NOTES
The quiescent centre in the root meristem serves as a reserve for replenishment of damaged cells of the meristem.
(3) Tunica corpus theory
- This theory was proposed by Schmidt (1924). This theory is applicable on shoot apex, it is based on planes of division.
- According to this theory, two types of layers are found in the shoot apex - Tunica & Corpus.
- Tunica : This is the peripheral layer. Epidermis is formed by this layer. In tunica cells, anticlinal division takes place only in one plane.

Anticlinal division occurs at right angles to the longitudinal axis of the cell. Surface area of the plant organs increases due to anticlinal divisions.
When division occurs in single anticlinal plane, they do not increase the number of layers.
Generally, tunica represents only a single layer, but sometimes it is multilayered, then the outermost layer forms the epidermis and remaining layers form rest types of the tissue system with the association of corpus.
- Corpus : The mass of cells present below the tunica is called corpus. The cells of corpus usually larger than the cells of tunica. It forms rest of the tissue system.
NOTES
The tunica and corpus of shoot apex is not distinguished in ferns and gymnosperm (except Gnetales).
PERMANENT TISSUES
- Permanent tissues are composed of cells which have lost the power of division temporarily or permanently. They are formed by division and differentiation of meristematic tissues.
- Cell cycle of permanent tissue is arrested at G1 stage so cells have no capacity to divide actively, so it is composed of mature cells.
NOTES
G1 stage of differentiated cell is called G0 stage.
- Their cells may be living or dead.
- Permanent tissues are of three types :
- Simple tissue (Homogenous tissue)
- Complex tissues (Heterogenous tissue)
- Special tissue
SIMPLE TISSUE
- These tissues are made up of similar cells that perform a common function.
- Simple tissues are of three types : parenchyma, collenchyma and sclerenchyma.
(A) PARENCHYMA
- It is very primitive type of tissue. It is the first evolved tissue. Remaining all different types of tissues are derived from this tissue. So it is also called as fundamental tissue.
- Characteristic features
- It is a living tissue and is the first to be differentiated from meristem.
- It is found in all plants and plant parts like root, stem and leaves.
- All the cells of parenchyma are thin walled and it contains both primary and secondary walls. Cell wall is made up of pectocellulose (mainly cellulose). So parenchyma is a soft tissue.
- Parenchyma is the most common tissue which is morphologically and physiologically unspecialized.
- Each cell contain large central vacuole. So the main function of a parenchyma cell is storage of food.
- Intercellular spaces are present between cells of this tissue.
- The cells are isodiametric. Flesh of a fruit is mainly composed of parenchyma.
- The cells of parenchyma are spherical, oval or polyhedral in shape.

- Modification of parenchyma
- Prosenchyma : The cells of this parenchyma are long with pointed ends. This parenchyma forms the pericycle of roots.
- Aerenchyma : This parenchyma is made up of rounded cells. These cells surrounds the large air chambers. It is found in cortex region. It provides buoyancy to hydrophyte plants.
- Chlorenchyma : In this parenchyma, abundant quantity of chloroplasts are found. Two types of chlorenchyma, present in dorsiventral leaves are :
- Palisade tissues : In this, intercellular spaces are absent. Number of chloroplasts are more in palisade tissue as compared to spongy tissue. So, upper surface of a leaf appears more green as compared to lower surface.
- Spongy tissues : Large intercellular spaces are present. So, they facilitate transpiration and gaseous exchange.
- Mucilage parenchyma : In the mucilage parenchyma, large vacuoles and mucilage will be found, e.g. Succulent xerophytic plants (Aloe) Function -storage of water.
- Functions of parenchyma
- The main function of this tissue is storage of food.
- Some cells of parenchyma store waste materials. They are called “idioblast cells”. Idioblast cells store and form oils, tannin and crystal of calcium oxalates.
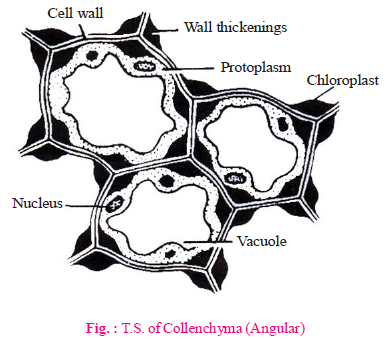
(B) COLLENCHYMA
- Characteristic features
- Collenchyma is a living mechanical tissue. Lignin is absent.
- It is made up of elongated, oval, spherical or polygonal cells.
- Localized deposition of pectocellulose (mainly pectin) & hemicellulose is the characteristic feature of collenchyma.
- Vacuolated cytoplasm is found in the cells of collenchyma.
- Collenchyma originates from ground meristem.
- Occurence
- Collenchyma is not a universal tissue. It is found in the stems of herbaceous dicotyledons.
- Collenchyma is absent in parts of woody plant parts, roots and monocotyledons.
- Collenchyma forms the hypodermis of dicotyledon stems. Cells of collenchyma are flexible due to hydrophilic nature of pectocellulose so flexibility occurs in dicotyledonous stems.
- Collenchyma is absent in plants after the secondary growth because plant becomes woody.
- Chloroplast may be found in the cells of collenchyma. Therefore, chloroplasts are found in the hypodermis of dicotyledonous stem.
- Functions
- Mechanical as well as biological. It provides tensile strength for extensibility of various growing plant organs.
- Due to the presence of chloroplast, it also participates in the process of photosynthesis.
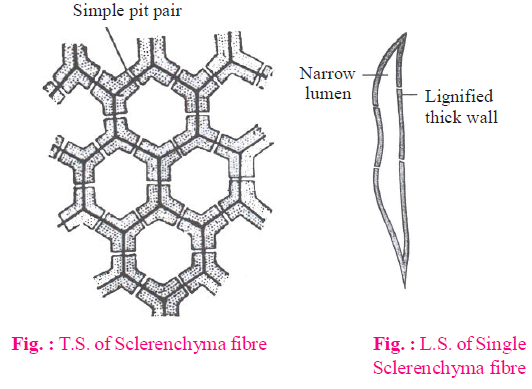
(C) SCLERENCHYMA
- Characteristic features
- Sclerenchyma is the main mechanical tissue.
- These cells are long, narrow, thick walled lignified and dead.
- Various types of pits are formed due to the deposition of lignin on hard wall.
- On the basis of length of cells and quantity of deposition of wall materials, sclerenchyma cells are of two types - sclereids and sclerenchymatous fibres.

- Sclereids
These cells are small, extremely thick walled and their ends are not pointed.
Sclereids are isodiametric or irregular in shape. Sclereids cells have more pits and lumen is almost very small. Their pit cavity is branched.
- Stone cells or Brachysclereids or Grit cells : These cells are spherical or oval in shape. They are found in endocarp of drupe fruits, so endocarp becomes hard.
They are present in endocarp of coconut, mango, almond, walnut, etc.
Besides drupe fruits, brachysclereids also present in fleshy edible part of pear. Grittiness in pear fruit is due to these sclereids.
- Trichosclereids : These are also known as internal hairs. They are spine like, bifurcated cells, found in floating leaves.
- Astrosclereids or Stellate sclerenchyma : These cells are stellate star shaped. They are found in floating leaves. E.g., Both Astro and Trichosclereids are present in floating leaves.
- Macro-sclereids or Rod cells or Malpighi cells : They are small and rod like cells. They are present in seed coats.
Example :
- They form a part of seed coat in legume plants. Due to their presence, seed coat becomes hard and dormancy is present in legume seeds.
- In leguminous plants, hardest seed coat is found in french bean. So, dormancy is found in leguminous seeds.
- In plant kingdom, hardest seed coat is found in lotus.
- Osteo-sclereids : These are known as prop cells. These are pillar like cells whose both end spreads to form bone like structure.
E.g., These cells are found in leaves of two plants Hakea and Osmanthus.
- Sclerenchymatous fibres
These cells are fibrous. They are the longest cells in the plant body. Their both ends are pointed tapering. Due to thick cell wall, lumen is reduced. Their cell wall contains simple and bordered pits.

- Xylary or wood fibres : These are hard fibres. The fibres are not flexible. They cannot be knitted (weaved) easily so they are not useful. These are found in xylem. e.g., Munj fibre (Saccharum munja)
- Bast fibres/Extra xylary fibres/Phloem fibre : These are also known as commercial fibres. These fibres are flexible and can be knitted easily. They have great economic value. These fibres are obtained from the phloem and pericycle of plants. The bast fibres of Corchorus capsularis (Jute), Crotalaria juncea (Sunn hemp) and Hibiscus sabdariffa (patua) are obtained from the secondary phloem of stem. The bast fibres of hemp (Cannabis sativa) and Linum usitatissimum (flax) are obtained from the pericycle. Fibres which are obtained from pericycle are called perivascular fibres.
IMPORTANT POINT
- Fibres are longest plant cell. Longest fibres occur in Boehmeria nivea (Ramie fibre), length: 55 cm.
- In plant kingdom, hardest seed coat is found in Nelumbo (Lotus).
- In plant kingdom, hardest, thickest and largest leaves are found in Victoria regia, due to astrosclereids. Diameter : 1-1.5 m.
- Longest leaves are found in Raphia vinifera. Length : 10-15 m.
COMPLEX PERMANENT TISSUE
- The complex tissues are made of more than one type of cells and these work as a unit. Complex tissue are heterogenous.
- Complex tissues are absent in gametophytes.
- Complex tissues are of two types - xylem and phloem. Xylem and phloem are together called conducting tissues organised into vascular bundles.
(A) XYLEM
- The function of xylem is to conduct water & minerals salts upwards from the roots to stem & leaves and to give mechanical strength to the plant body.
- On the basis of origin, xylem is divided into primary xylem and secondary xylem.
- Primary xylem originates from procambium. On the basis of development, primary xylem is divided into two parts- protoxylem & metaxylem. Cells of protoxylem are small as compared to metaxylem. Secondary xylem originates from vascular cambium.
- The elements of xylem are-

(1) Tracheids
- Tracheids are primitive conducting elements of xylem.
- Tracheids are unicellular and having narrow lumen (but lumen of tracheids is wide than fibres).
- Tracheids join together from their ends to form long rows. These rows extending from the roots via stem to the leaves.
- A transverse septum lies between each two tracheids. Water moves from one tracheid to another tracheid through pits.
- Tracheids are dead and lignified cells. The deposition of lignin on cell wall is responsible to form a different type of thickenings (Pits are unlignified areas on lignified wall).
NOTES
- Usually bordered pits are present at end wall of tracheids.
The maximum bordered pits are found in the tracheids of Gymnosperm plants.
- Maximum deposition of lignin is found in pitted type of thickening.
- Annular and Spiral type of thickening of lignin is found in protoxylem.
- Reticulate and Pitted (mainly) type of thickening of lignin is found in metaxylem.
- Tracheids of pteridophytes have long or elongated bordered pits. Such type of pits are called scalariform pits.
(2) Vessels
- It is an advance conducting element of xylem. The basic structure of vessels is same as tracheids.
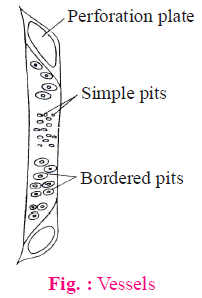
- Vessel are multicellular. These are also dead elements of xylem.
- The lumen of vessels is wider than tracheids and end wall is perforated. Transverse septum is absent between two vessel elements, if present then porous. Thus, vessels are more capable for conduction of water than tracheids. Due to presence of perforated end wall, vessels work as a pipeline during conduction of water.
- Vessels contain usually simple pits at their lateral walls. Thickening type of wall is the same as tracheids.
NOTES
- Vessels are only found in xylem of angiosperm but exceptionally it is also present in some Gymnosperms like Ephedra, Gnetum and Welwitschia.
- Vessels are absent in some angiospermic plants such as Dracaena, Yucca, Dazinaria, Drimys. There are some angiosperm families in which vesselless angiosperms are included, e.g., winteraceae, tetracentraceae and trochodendraceae.
- Vessels are examples of dead syncyte. Cell which is formed by fusion of cells is called as syncyte.
(3) Xylem fibres
- These are also dead parts of xylem . Xylem fibres provides strength to the tracheids and vessels. Mainly these fibres provide strength to the vessels.
- They are present more abundant in secondary xylem.
(4) Xylem parenchyma
- Only xylem parenchyma is a living cell of xylem.
- Its cell wall is made up of cellulose. It stores starch, fats and tannin etc.
- The radial conduction of water is the function of xylem parenchyma. (It conducts water to peripheral part of plant organ).
- Their walls possess pits.
(B) PHLOEM
- Phloem is called bast or leptome. The main function of the phloem is to conduct food materials, usually from the leaf to other plant parts (e.g., storage organ and growing regions).
- On the basis of origin, phloem is classified into two categories - primary and secondary phloem.
- Primary phloem originates from procambium and secondary phloem originates from vascular cambium.
- On the basis of development, primary phloem is categorised into protophloem and metaphloem.
- The protophloem has narrow sieve tubes whereas metaphloem has bigger sieve tubes.
- Phloem remains active for less duration as compared to xylem.
- Phloem consists of the 4 types of cells : Sieve elements, companion cells, phloem parenchyma and phloem fibres.
(1) Sieve elements
- Sieve element was discovered by Hartig. Sieve elements includes sieve cells and sieve tubes.
- Sieve cells/sieve tube element are living and thin walled.
- A matured sieve cell/sieve tube elements lack nucleus. Thus, these are enucleated living cells.
- Central vacuole is present in each sieve cell/sieve tube element. Cytoplasm of sieve cells/sieve tube element show cyclosis in the form of thin layer around the vacuole.
- In angiosperm plants, sieve tube elements are arranged with their ends and form sieve tube.
- Sieve plate (oblique transverse perforated septa) is present between the two sieve tube elements at their end wall. It is porous. Materials are transported through these pores.
- Callose β-1-3 glucan deposited on the radius of pores during dropping season (autumn) of leaves, to form a thick layer. This is called callus pad.
- Sieve plate is protected by callus pad. It is also prevented from bacterial infection and drought.
- Callose dissolves during spring season.
- In gymnosperms and pteridophytes, sieve cells do not form sieve tube and arranged irregularly. Sieve cell have sieve plates on their lateral walls. Thus, conduction of food takes place in zig-zag manner.
- In angiosperms, food conduction is erect and efficient.

NOTES
- Food conduction is bidirectional in sieve tube.
- In sieve tube, oblique transverse perforated septa are present on their end wall.
- Sieve tube is an example of living syncyte.
(2) Companion cells
- Companion cells are thin walled living cell with large nucleus. This nucleus also controls the cytoplasm of sieve tube element.
- Companion cells are only found in angiosperms.
Exception - Austrobaileya is angiosperm plant but companion cells are absent.
- The sieve tube elements and companion cells are connected by pit fields present in their longitudinal walls, which is a common wall for both and, with the death of one, other cell also dies.
- A companion cell is laterally associated with each sieve tube element in angiospermic plants.(ln carrot, it is more than one).
- Sieve tube element and companion cell originates together. Both of them originates from a single mother cell. So, called as sister cells.
- The companion cells and sieve tube elements maintain close cytoplasmic connections with each other through plasmodesmata.
- Special type of cells are attached with the sieve cells in gymnosperms and pteridophytes in place of companion cell. These cells are called albuminous cells/strasburger cell.
NOTES
- Albuminous cells in conifers are analogous to companion cells of angiosperms. They are modified phloem parenchyma cells.
- The companion cells play an important role in the maintenance of a pressure gradient in the sieve tubes.
- In phloem of pteridophytes and gymnosperms, sieve cells are present which are comparable to the tracheids.
(3) Phloem fibres
- Phloem fibres may be living or nonliving.
- Living phloem fibres are useful for storage of food materials while non-living fibres provide mechanical support to the conducting elements (sieve cells and sieve tube). Main function of these fibres are providing mechanical support and can be used in making ropes, thick clothes and mats.
(4) Phloem parenchyma
- It is also known as bast parenchyma.
- Its cells are living and thin walled. It stores various materials, e.g., resin, latex, mucilage etc.
- The main function of phloem parenchyma is conduction of food in radial direction and storage of food.
NOTES
- Phloem parenchyma is absent in the stems of monocotyledon plants.
- Phloem parenchyma is absent in the stems of Ranunculaceae plants (dicot family), e.g., Thalictrum.
SPECIAL TISSUE OR SECRETORY TISSUE
(A) LATICIFEROUS TISSUE
- These are made up of long, highly branched and thin walled cells. These cells are filled with milky juice, called as latex.
- Latex is the mixture of saccharides, starch granules, alkaloids, minerals and waste materials.
- Function
- Latex provide protection to the plant.
- It prevents the plants from infection of bacteria and fungus. In laticiferous tissue, there are two types of cells- latex vessels and latex cells.
- Latex vessels : These are articulated vessels and formed due to dissolution of cells walls of meristematic cells. Thus, these are syncytic cells as well as coenocytic (multinucleated). E.g., Latex vessels are present in Hevea, Ficus, Papaver, papaya, Argemone and Sonchus.

Highly developed latex vessels are found in the fruit wall of Poppy.
Opium is obtained from the latex of Papaver somniferum. It contains an alkaloid named as morphine.
An enzyme papain is obtained from the latex of papaya (Carica papaya).
Indian rubber is obtained from Ficus elastica and para rubber is obtained from Hevea brasiliensis.
Mostly latex is white in colour but in some plants, latex is coloured.
E.g., Papaver is dark brown and Argemone and Sonchus are in yellow colour.
In some plants, latex is colourless, E.g., Banana.
- Latex-cells : These are non-articulated latex ducts/tubes. These are long, branched and multinucleated cells (coenocytic cells).
E.g., Latex cells are found in Calotropis, Euphorbia and Nerium, Ficus religiosa
(B) GLANDULAR TISSUE
- This tissue is made up of glands which contain secretory or excretory materials.
- Glandular tissues have two types of glands
(1) Unicellular
These cells are present on the surface of the leaves. These are spiny glands in which formic acid is filled. It protects the plants from grazing animals.
E.g., Urtica-dioica.
E.g., Urtica-dioica.
(2) Multicellular
Multicellular glands are of two types:
- External glands : These are located on the surface of the plants and arising as an outgrowth from the epidermis. These glands are of various types-
- Glandular hairs : They secrete gum-like sticky substances in tobacco and Plumbago, digestive juicy substance in Drosera.
- Nectar glands : These glands secrete sugar solution. These are found in floral parts mainly in thalamus. These glands secrete nectar to attract the insects.
Exception : In Passiflora, nectar glands are found in leaves.
- Internal Glands : These glands are embedded in the tissues. Internal glands are of following types:
- Digestive glands : Digestive glands are found in insectivorous plants. These glands compensate the deficiency of N2 in insectivorous plants. These are found in Utricularia, Drosera, Dionaea etc. plants. They secrete proteolytic juice.
- Mucous secreting glands : These glands secretes mucus. These are found in the leaves of betel.
- Oil glands : These secrete volatile oil. It acts as an antiseptic. These glands are found in fruits & leaves of lemon & orange. Mostly, oil glands are lysigenous but in sunflower, these glands are schizogenous.
NOTES
- Oil glands which secrete volatile oils called osmophores. Osmophores develop fragrance in flowers.
- Oils which are obtained from Eucalyptus leaves are used in medicines.
- Tannin, resin, gum, secretory glands are also internal glands.
- Maximum resin glands are found in Pinus.
- Resin ducts are schizogenous. Gum glands are found in Acacia.
TISSUE SYSTEM
- In higher plants, several tissues work together in the form of a unit to perform a particular function. These tissues have the same origin. Such tissues form a system which is called tissue system.
- On the basis of division of labour, tissue is categorised by Sachs into three different system - epidermal tissue system, ground tissue system and vascular tissue system.
- Each system usually consists of an association of tissues which perform specific function.
EPIDERMAL TISSUE SYSTEM
- This system includes epidermis and its related structures, e.g., trichomes, root hairs, stomata and bulliform cells etc.
- Epidermis is the outermost layer of all organs of primary plant body.
- All the epidermal cells are living (parenchymatous) & contain vacuolated protoplasm.
- The epidermis is covered with a cutin layer called cuticle on its outer surface.
- The thickness of cuticle depends upon the adaptation of plant. It is thick in xerophytes, thin in mesophytes & absent in hydrophytes.
- Cuticle prevents the less of water and it is absent in roots.
- The cells of the epidermis gives rise to a number of protuberances which vary markedly in their shape, structure and function. These are known as trichomes. These appendages can be unicellular or multicellular. The unicellular trichomes are usually simple, unbranched (sometimes also branched) and elongated structures. On the other hand, the multicellular trichomes and glands are made up of several layers of cells. In shoot system, trichomes are usually multicellular.
- The trichomes help in protection, dispersal of seeds and fruits, and reduction of water loss.
ROOT HAIRS
- The root hairs are formed due to the elongation of the epidermal cells and are not protuberances or appendages. These have a vacuolated protoplasm and the nucleus moves towards the apical part of the cell. The thin wall is made up of cellulose and pectic materials.
- Root hairs play an important role in anchoring the plant body in the soil besides absorbing water and mineral solution from it.
STOMATA
- Stomata are tiny pores. Stomata are composed of two bean shaped epidermal cells called guard cells which encloses stomatal pore.
- Stomata are mostly present in epidermal layers of leaves and in other aerial parts like young stems, floral parts etc.
- Stomata are absent in roots.
- Guard cells in dicots are kidney shaped and in monocots are dumb bell shaped.
- Outer wall of guard cell is thin but inner wall is thick & inelastic.
- The movement of stomata depends on the turgidity & the flaccidity of guard cells.
- In majority of plants, stomata open during day time and remain closed during night.
- The guard cells may be surrounded by varying number of specialized epidermal cells called subsidiary cells or accessory cells.
- The stomatal aperture, guard cells and the surrounding subsidiary cells are together stomatal apparatus.
- Having stomata, epidermis helps in interchange of gases and transpiration.
GROUND TISSUE SYSTEM
TRICHOMES
- Ground tissue system, also known as fundamental tissue, contain various types of tissue excluding epidermis and vascular tissue.
- It is the largest tissue system.
- Ground tissue system is derived partly from periblem and partly from plerome.
- It includes, general cortex, hypodermis, endodermis, pericycle and medullary rays (pith rays).
- Cortex is the zone which lies between the epidermis & the pericycle.
- Hypodermis in stems & leaves in dicot is collenchymatous while it is sclerenchymatous in monocots.
- In stems and leaves, hypodermis gives mechanical support. In dicot stem, hypodermis performs photosynthesis (when it contains chloroplast) and give rise to cork cambium.
- Endodermis is the innermost layer of cortex.
- Endodermal cells are living and contain abundant protoplasm and large nuclei.
- Endodermal cells of roots are characterised by casparian strips.
- Pericycle is single or few layered parenchymatous or rarely sclerenchymatous cells. In angiosperms, it gives rise to lateral roots.
- Pericycle is absent in the stems of monocot, hydrophytes and parasites.
- Main function :
- Manufacture and storage of food material.
- Mechanical support (additional work)
VASCULAR/STELE/CONDUCTING TISSUE SYSTEM
- This tissue system originates from procambium.
- It consists of xylem and phloem. It is also called specific tissue system.
- Phloem elements are formed earlier than the xylem elements.
WATER CONDUCTING ELEMENTS OF XYLEM
Types of development of conducting elements are centrifugal, centripetal and both.
- Centrifugal : In this type of development, the protoxylem formed near the central axis and metaxylem is formed away from the centre it means towards the periphery. This condition is known as endarch. E.g., stem of angiosperm and gymnosperm
- Centripetal : In this, protoxylem is formed away from the centre it means near the pericycle and metaxylem is formed towards the centre. This condition is called exarch. E.g., Roots.
- Centrifugal and Centripetal : In this, elements of metaxylem is formed from both sides of the elements of protoxylem. In this type of development, protoxylem is surrounded by metaxylem. This condition is known as mesarch. E.g., Fern rhizome.
VASCULAR BUNDLES
- Xylem and phloem are collectively termed as vascular bundles.
- On the basis of arrangement of different parts, vascular bundles are divided into three categories; radial vascular bundles, conjoint vascular bundles and concentric vascular bundles.
(A) Radial vascular bundles
When the xylem and phloem are present separately on different radii in alternate manner, it is called radial vascular bundles. The order of development of xylem in these vascular bundle is centripetal. Thus, these vascular bundles are called exarch.
E.g., radial vascular bundles is always closed and present in most of the roots.

Exception : Radish, carrot, turnip, sugar beet/Beetroot/Beta vulgaris (conjoint-collateral)
(B) Conjoint vascular bundles
- In this type of vascular bundle, xylem and phloem are present on the same radius.
- These are of two types - conjoint collateral and conjoint bicollateral and open vascular bundles.
(1) Conjoint Collateral
In conjoint collateral vascular bundle, xylem and phloem are present on the same radius and phloem present towards the periphery.

These are two types - open and close.
- Open : If the cambium is present between the xylem and phloem, it is said to be open vascular bundle. E.g., stem of dicotyledons and gymnosperms.
- Close : When cambium is absent between the xylem and phloem, in conjoint vascular bundle, it is called closed vascular bundle. E.g., monocotyledons stem.
In this type of vascular bundle, order of development of xylem is centrifugal. So, endarch condition is found in xylem.
(2) Conjoint bicollateral and Open vascular bundle
There are two patches of phloem, one on each side of xylem, are found. There are two strips of cambium (outer and inner), one on each side of xylem. Such type of vascular bundles are known as conjoint, bicollateral and open vascular bundle. Only outer cambium is functional.
Order of development of xylem is centrifugal so endarch condition is found.
E.g., Stem of family cucurbitaceae (Luffa cylindrica, Cucurbita pepo & Lagenaria stem) and some plants of family Apocynaceae and Solanaceae.
(C) Concentric vascular bundles
In this type of vascular bundle, either xylem surrounds the phloem or phloem surrounds the xylem. Concentric vascular bundles are always closed. They are of two types - amphicribral and amphivasal.
(1) Amphicribral or Hadrocentric
- In this type of vascular bundle, xylem is completely surrounded by phloem. It means xylem is present in the centre of vascular bundle.

- The order of development of xylem in these vascular bundles are both in centripetal and centrifugal manner. In this type of vascular bundle, protoxylem is surrounded by metaxylem. These are known as mesarch vascular bundle.
- Such types of vascular bundles are found in stem of ferns and lower gymnosperms.
(2) Amphivasal or Leptocentric
- In this type of vascular bundle, phloem is completely surrounded by xylem. It means phloem is present in the centre of the vascular bundle.
- In this type of vascular bundle, xylem is endarch, e.g., stem of Dracaena, Yucca etc.
STELE
- The stele is the whole central mass of vascular tissue (vascular cylinder) with or without pith surrounded by endodermis.
- Van tieghem and Douliot put forward the hypothesis about stele. According to him, stele is the central part or core of the axis of the plant which includes the vascular system and its related structures.
- The tissues which lies inside the stele is called intrastelar tissues and the tissues which lies outside the stele is known as extrastellar tissues.
- Stele is surrounded by endodermis but endodermis is originally the part of cortex. It is not a part of stele.
- Stelar system started from pteridophytes.
- In order of evolution, types of stele are - protostele, siphonostele, solenostele, dictyostele, eustele and atactostele.
(A) Protostele or Monostele or solid stele
- Protostele is the most primitive and simplest type of stele.
- It consists of a solid mass of xylem which is completely surrounded by phloem. Such type of stele lack pith.
- Solid stele is of following types (on the basis of shape of xylem) :
- Haplostele : In this stele, xylem is surrounded by a smooth layer of phloem. Central xylem is cylindrical but circular in T.S. E.g., Rhynia, Selaginella selaginoides etc.
- Actinostele : Actinostele is that stele in which the central xylem has radiating ribs and assumed a star shaped appearance. E.g., Psilotum and lsoetes.
- Plectostele : In this solid stele, the xylem divides into number of separate plates which lie parallel to one another. E.g., Most of the species of Lycopodium.
- Mixed protostele : Sometimes the solid xylem core of the protostele is broken into small group of tracheids which remain embedded in the phloem. Such a protostele is known as mixed protostele. E.g., Lycopodium cernum.
(B) Siphonostele
- Siphonostele is that stele in which the pith is present in the centre of vascular cylinder.
- Siphonostele is of two types :
- Ectophloic siphonostele : In vascular tissue of such type of stele, phloem is always present outside of the xylem. E.g., Equisetum, Osmunda
- Amphiphloic siphonostele : In vascular tissue of such type of stele, xylem is surrounded by phloem on both sides. E.g., Adiantum, Marsilea
(C) Solenostele
- In pteridophytes, there occur two types of leaves - microphyllous and megaphyllous.
- Megaphyllous leaves require more vascular supply and these leaves receive vascular supply from stem. When a megaphyllous leaf develops on stem, vascular cylinder of plant organ (stem) breaks from one side and a gap is formed. It is called as leaf gap (xylem & phloem are absent in leaf gap and it is filled with parenchyma).
- Vascular supply diverting from stem to leaf is called leaf trace. (Leaf trace and leaf gap are present in T.S. of stem and opposite to each other at internode region). Due to the formation of one leaf gap, stele becomes horse shoe shaped called solenostele.
- Solenostele also may be ectophloic or amphiphloic.
(D) Dictyostele or polystele
- When the production of many leaf gaps in siphonostele, main vascular cylinder breaks into many fragments, then such type of siphonostele is called dictyostele.
- Each divided fragment (piece) is called meristele. Each meristele has its own separate endodermis and pericycle.
- Pith is absent in meristele. Meristele is complete stele so dictyostele is well developed type of stele in pteridophytes. E.g., Pteridium, Pteris, Dryopteris
(E) Eustele
- In this type of stele, vascular bundle are arranged in a ring and medullary rays are present between vascular bundle. Such type of stele is found in stem of gymnosperm and dicotyledon plants.
- Normally in Cycas stem, eustele is present but in Cycas young stem vascular cylinder is very small in comparison to the cortex and several vascular bundles are arranged in a ring forming an ectophloic siphonostele.
(6) Atactostele
- Many vascular bundles are distributed in ground tissue. Such type of stele is called atactostele.
- Endodermis and pericycle are absent in atactostele. This is highly developed type of stele.
- Such type of stele is the main characteristic feature of monocotyledons.



INTERNAL STRUCTURE OF STEMS, ROOTS AND LEAVES
INTERNAL STRUCTURE OF DICOT STEMS
Internal structure of a typical dicot stem shows following features like epidermis, cortex, pericycle etc.
EPIDERMIS
- Epidermis is the outermost layer of the stem. It is single layered and lack chloroplast.
- Multicellular hairs (trichomes) and stomata are found on epidermis.
- Outer side of epidermis a layer is present which is made up of cutin is called cuticle.
- Epidermis plays a significant role in protection.
CORTEX
- In dicotyledon stem, cortex is divided into three parts - hypodermis, general cortex and endodermis.
- Hypodermis is present just below the epidermis and provides additional support to epidermis. It is thick multicellular layer, composed of collenchyma and their cells contain chloroplast. So, hypodermis is green and photosynthetic.
- General cortex is composed of parenchyma. Storage of food is the main function of the cortex. Resin canal/mucilage canal are present in it. These are schizogenous in origin.
- Endodermis is single celled thick layer. The innermost layer of the cortex is called endodermis. The cells of endodermis are barrel shaped. These cells accumulate more starch in stem of dicot. Thus, it is known as "starch sheath".
PERICYCLE
- This layer is situated in between the endodermis and vascular bundles.
- The pericycle of stem is multilayered and made up of sclerenchyma.
- Sclerenchymatous pericycle is also known as hard bast.
NOTES
In sunflower stem, pericycle is made of alternate bands of parenchymatous and sclerenchymatous cells. Pericycle which is present in front of the vascular bundle is made up of sclerenchyma and remaining is composed of parenchyma. Part of pericycle which is situated in front of vascular bundle is known as bundle cap. Pericycle is heterogenous in sunflower stem.
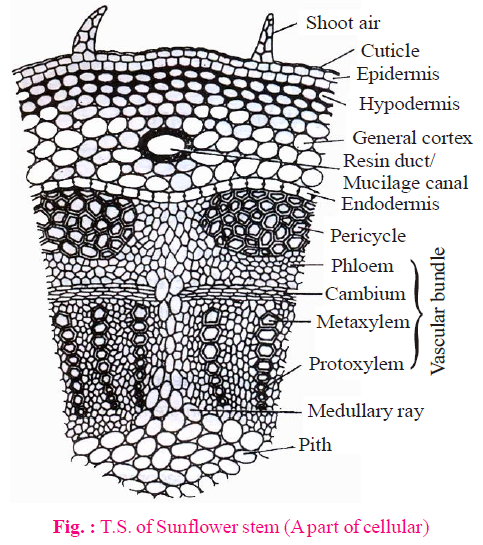
VASCULAR BUNDLE
- The vascular bundles (wedge shaped) are arranged in a ring.
- Each vascular bund is conjoint, collateral and open.
- Each vascular bundle is made of phloem, cambium and xylem.
- Eustele is present in dicotyledon stems.
PITH
- This is well developed region, spreading from ring of vascular bundle to the centre.
- The cells of this region mainly made up of parenchyma.
- Function of pith is storage of water and food.
NOTES
The part of pith which is radially arranged between the vascular bundles, called pith rays or medullary rays. The main function of pith rays is radial conduction of food and water.
E.g., Cucurbita stem:
- It contains five ridges and five furrows.
The vascular bundles are arranged in two rows. Each ring has five vascular bundles. In this way, the total 10 vascular bundles are present.
- The vascular bundles of outer ring are small in size and situated in front of ridges while the vascular bundles of inner rings are large in size and located below the furrows.
- Vascular bundles are conjoint, bicollateral and open.
- Hypodermis is absent or less developed in furrow region and cortex contains chloroplast.


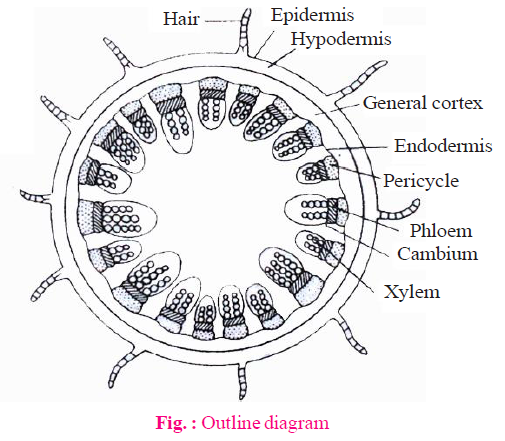
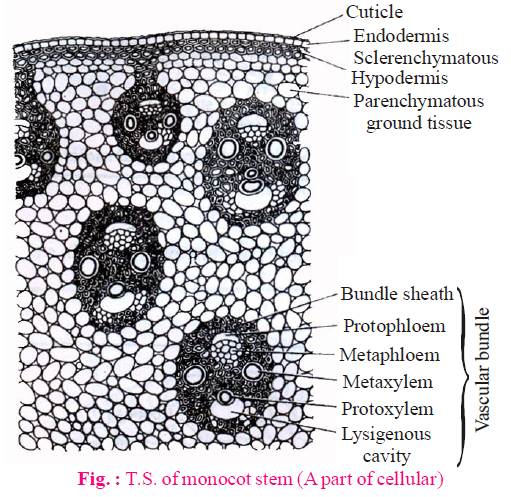
INTERNAL STRUCTURE OF MONOCOT STEM
EPIDERMIS
- Epidermis is the outermost single celled thick layer.
- It is covered with thick cuticle. Multicellular hairs are absent and stomata are also less.
HYPODERMIS
- Hypodermis of monocotyledon stem is made up of sclerenchyma. It is 2-3 layered.
- In monocot stem, rigidity is more in hypodermis whereas in dicot stem elasticity is more.
- It provides mechanical support to plant.
GROUND TISSUE
- The entire mass of parenchyma cells next to hypodermis and extending to the centre is called ground tissue.
- There is no differentiation of ground tissue in monocotyledon stem. It means ground tissue is not differentiated into endodermis, cortex, pericycle etc.
NOTES
Sometimes in some grasses, wheat etc. the central portion of ground tissue becomes hollow and is called pith cavity.
VASCULAR BUNDLE
- Many vascular bundles are scattered in the ground tissue and they are generally oval shape.
- Vascular bundles lies towards the centre are large in size and less in number.
- Vascular bundles situated towards the periphery are small in size but more in number.
- Each vascular bundles are conjoint, collateral and closed.
- Vascular bundles surrounded by the layer of sclerenchymatous fibre is known as bundle sheath. So, vascular bundles are called fibrovascular bundles.
- Xylem : In xylem, number of vessels is less. In metaxylem, there occur two large vessels while in protoxylem, there occur one or two small vessels. Vessels are arranged in V or Y shape. Just beneath protoxylem vessels, there occur a water cavity which is schizolysigenous in origin but major part of water cavity is lysigenous. This cavity is formed by disintegration of the element present below the protoxylem and neighbouring parenchyma.
Exception : In Asparagus, water cavity & bundle sheath are absent.
- Phloem : It consists of sieve tube elements and companion cells. Phloem parenchyma is absent.
PITH
- Pith is undifferentiated in monocotyledon stems.
- Atactostele is found in monocotyledon. This is highly developed stele.


INTERNAL STRUCTURE OF DICOT-ROOT
Internal structure of a typical dicotyledon root shows following features like epiblema, cortex, endodermis etc.
EPIBLEMA
- It is uniseriate outermost layer. It comprises of tubular living components.
- Cuticle and stomata are absent.
- Unicellular root hairs are formed due to elongation of some cells of epiblema.
- Epiblema is also known as rhizodermis or piliferous layer. Root hairs are present in maturation zone of root.
- Cells of epiblema which develop root hair called trichoblast.
CORTEX
- It is made up of parenchymatous cells.
- Chloroplast is absent so they are non-photosynthetic but chloroplast is present in roots of Tinospora and Trapa so they are photosynthetic.
- The cells of outer part of cortex are suberized in old root. It is called exodermis.
- Exodermis is found in some dicotyledonae roots and most of the monocotyledonae roots.
ENDODERMIS
- This layer is situated between the pericycle and cortex.
- Casparian strips are present on radial and tangential wall of endodermis. These strips are made up of lignosuberin (mainly suberin).
- The cells of endodermis which are situated in front of protoxylem cells, lack casparian strips. These are called passage cells/ transfusion cells.
- The number of passage cells is equivalent to the protoxylem cells. Passage cells provide path to absorbed water from cortex to pericycle.

- Root hairs are linearly arranged on root apex. Number of root hairs are similar to the number of passage cells.
- Casparian bands and passage cells are well developed in monocot root. They are also present in stem but they are less developed.
- Endodermis acts as a watertight jacket.


PERICYCLE
- It is single layered and composed of prosenchyma, a type of parenchyma.
- Pericycle of roots are single layered. At the time of secondary growth, pericycle is responsible for the development of cork cambium.
- Lateral roots are originated from the part of pericycle which is lying opposite to protoxylem. Thus, lateral roots are endogenous in origin.A few mature cells of pericycle usually opposite to protoxylem, become meristematic. These cells divide by periclinal divisions and form some layers of cells. These divisions are followed by anticlinal divisions forming a primordium which grows to form a lateral root.
- The branches of stems are exogenous in origin because these originate from the outer part of cortex.
- Adventitious roots are also endogenous. It originates from phloem parenchyma or internal layer of endodermis or pericycle.
VASCULAR BUNDLES
- Vascular bundles are radial and exarch. Xylem and phloem are separate and equal in number.
- The number of xylem bundles are two to six (diarch to hexarch).
But exceptionally, Ficus (Banyan tree) root is polyarch.
- Parenchyma which is found in between xylem and phloem is called conjunctive tissue.
PITH
In dicot root, pith is less developed or absent.
INTERNAL STRUCTURE OF MONOCOT ROOT
- The internal structure of a typical monocotyledon root is similar to dicotyledon root.
- Number of xylem bundles are more than six (Polyarch) in monocotyledon root (exceptionally the number of xylem bundles are two to six in onion).
- Pith is well developed in monocotyledon root.


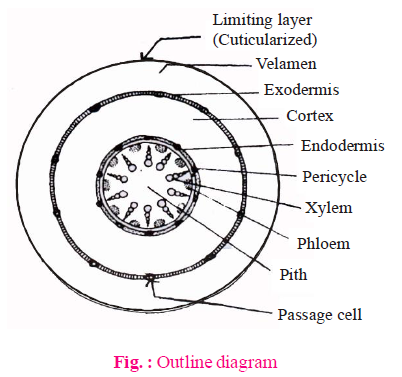
INTERNAL STRUCTURE OF ORCHID ROOT
- Shows the following features
- Velamen : These are found in aerial or hanging roots of some epiphytes (e.g., orchid).
- These are examples of multilayered epidermis. These are present outside the exodermis.
- These are made up of protoderm and are dead.
- They absorb atmospheric moisture by imbibition.
- Passage cells are found in both exodermis and endodermis in hanging roots of orchids.

INTERNAL STRUCTURE OF LEAF
Generally leaves divided into two categories - dorsiventral leaves and isobilateral leaves.

INTERNAL STRUCTURE OF DORSIVENTRAL LEAVES
- Dicot leaf is characterized by upper and lower epidermis, cuticle, mesophyll cells and vascular bundles.
- Cuticle is present on both surfaces but cuticle of upper surface is thicker.
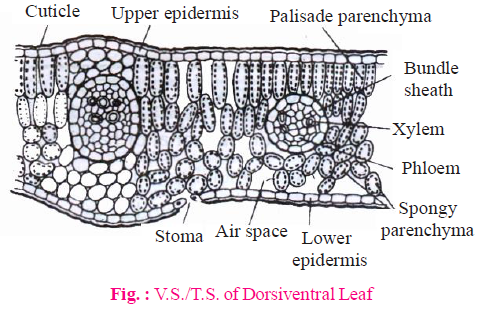
- Dorsiventral leaves are mostly hypostomatic i.e. stomata present on lower surface.
- In amphistomatic dorsiventral leaves, stomata are more on lower surface as compared to upper surface.
- Mesophyll of these leaves is divided into two regions - palisade tissue and spongy tissue.
- Palisade tissue is found towards upper surface. These cells have more chloroplasts and spongy tissue is found towards lower surface.
INTERNAL STRUCTURE OF ISOBILATERAL LEAVES
- The thickness of cuticle on the both surface is equal.
- Distribution of stomata on both surface are equal.
- Isobilateral leaves are amphistomatic i.e. stomata present on both sides.
- Both the surface of isobilateral leaves receives equal amount of sunlight. Thus, the structure of both the layers are the same.
- Mesophyll of isobilateral leaves is not differentiated into palisade and spongy tissues. It is completely made up of spongy tissues. Palisade tissue are absent.
- Bulliform cells or motor cells are present.
- Bulliform cells are large cells found in the epidermis of psammophyte (desert) grasses which are filled by liquid or empty (mostly) and colourless. Upward folding of leaves during the sunny day takes place due to the presence of these specific cells. This is an adaptation to reduce the transpiration.
E.g., Ammophila, Poa, Empectra and Agropyron etc. are Psammophytic grasses.
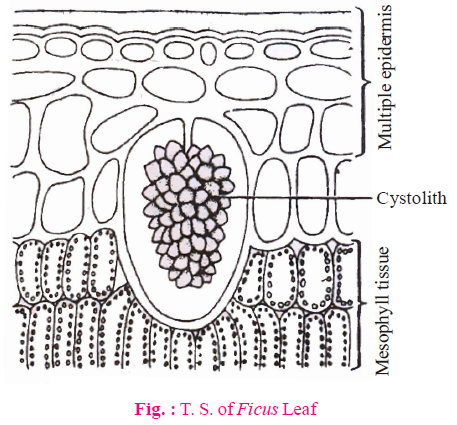
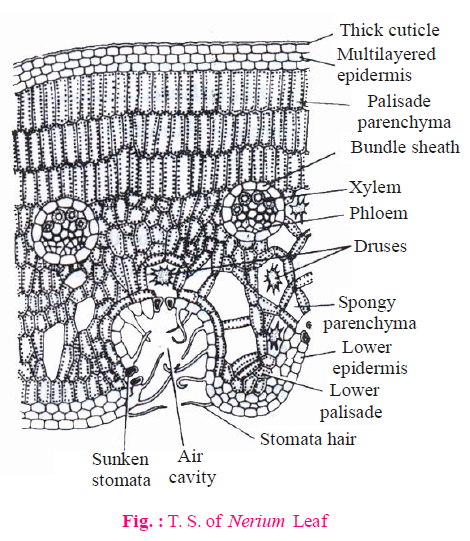
- Epidermis of Nerium (both upper & lower) and Ficus (only upper epidermis) becomes multilayered. This is an adaptation to reduce transpiration.

NOTES
- In isobilateral leaf, two distinct patches of sclerenchyma are present above and below each of the large vascular bundles and extend up to the upper and lower epidermal layers, respectively.
- In dorsiventral leaf, two distinct patches of parenchyma/sclerenchyma are present above and below each of the large vascular bundles and extend up to the upper and lower epidermal layers, respectively.
VASCULAR BUNDLES OF LEAVES
- Similar types of vascular bundles are found in both dorsiventral and isobilateral leaves. Vascular bundles of leaves are conjoint, collateral and closed.
- Protoxylem is situated towards the adaxial surface and protophloem towards the abaxial surface in the vascular bundle. Leaves are devoid of endodermis and pericycle.
- Vascular bundles are surrounded by a bundle sheath. Bundle sheath in C-4 plants is chlorenchymatous and remaining plants have parenchymatous or sclerenchymatous (mainly parenchymatous) bundle sheath.
IMPORTANT FACTS
- Xerophytes with isobilateral leaves contain palisade tissue on both sides of the leaf.
- Desert grasses contain palisade like spongy tissue.
- Unifacial or cylindrical leaf have no differentiation of upper surface and lower surface. E.g., Onion, Garlic.
- Albascent leaf : Palisade tissue is restricted in half part of leaf so half part appears more green and other half appears less green. E.g., Abutilon
- Tetrarch Condition → Helianthus annuus and Cicer arietinum root
- Waiting meristem concept : This concept was given by Buvat. According to this, there is an inactive centre in the shoot apex which is known as waiting meristem and it acts as a reservoir of active initials and on induction it give rise to reproductive apex.
- Tannin is found in latex of banana. When it comes in contact with air, it gets oxidized and becomes reddish brown in colour.
- Tannin glands are found in Camalia. These glands are schizogenous in origin.
- Salt glands are found in Tamarix which secretes sodium chloride.
- Chalk glands are found in plants of plumbaginaceae family which secretes calcium carbonate.
- Cystolith cells found in upper epidermis of Ficus leaf are called lithocysts. Cystolith are crystals of CaCO3.
- The most durable wood is Tectona grandis.
- Tracheids are the chief water transporting elements in gymnosperms.
- Phloem is embedded into the secondary xylem in some plants. Such phloem is called included phloem or interxylary phloem. This is secondary anomalous structure.
E.g., Leptadaenia, Salvadora etc. dicot stem.
- In some monocot roots, pith is sclerenchymatous. E.g., Canna.
- In aquatic plants and ferns, the epidermal cells and guard cells of stomata possess chloroplasts.
SECONDARY GROWTH
- Cells of apical meristems divide, differentiate and develop to form primary tissues. As a result, the plant grows in length. This is called primary growth.
- By the activity of secondary lateral meristems, increase in the circumference/girth of the plant organs due to the formation of secondary tissues in stelar & extrastellar regions, is called secondary growth.
- Normally secondary growth takes place in roots and stems of dicotyledons & gymnosperms.
- Due to lack of cambium in monocotyledons, secondary growth is absent. But exceptionally secondary growth takes place in some monocotyledons, such as - Palm, Yucca, Dracaena, Smilax, agave, coconut etc.
SECONDARY GROWTH IN DICOT STEM
(A) SECONDARY GROWTH IN STELLAR REGION
Secondary growth in stelar region begins earlier than the extra stelar region.
(1) Formation of ring of vascular cambium
- A cambium which is present inside the vascular bundle is called intrafascicular cambium. This is a type of primary meristem.
- First of all, cells of medullary rays become meristematic to form interfascicular cambium which is secondary lateral meristem.
- Intrafascicular and interfascicular cambium are collectively known as vascular cambium (VC). Vascular cambium is formed in the form of a complete ring which is made up of single layer of cells.
- In dicot stem, some part of V.C. is primary and some part is secondary.
- Two types of cells are found in the ring of this vascular cambium - fusiform initials and ray initials.
- Fusiform initials are long with pointed ends, while ray initials are spherica1 i.e., oval.
- Amount of fusiform initials is more in vascular cambium.
(2) Activity of vascular cambium
(a) Activity of fusiform initials
- Continuous periclinal divisions or tangential division takes place in fusiform initials.
- The plane of division in periclinal divisions is parallel to longitudinal axis of a cell. Through this type of activity, few cells are formed towards the radius (periphery) and these cells differentiate into secondary phloem or bast and some of the cells are formed towards the central axis and these cells are differentiated into secondary xylem or wood.
- Normally more secondary xylem is formed as compared to secondary phloem due to unequal distribution of hormones.
(Secondary xylem is formed 8-10 times more as compared to secondary phloem).
By the pressure of secondary phloem, primary phloem is pushed towards the outside and gets crushed.
- By the pressure of secondary xylem, all the primary tissues- such as primary xylem, pith etc. and old secondary xylem degenerates in the centre of the stem. Due to this, central part of the stem becomes woody. These activities keeps going on continuously in plants throughout the life.



- Secondary xylem forms in the plant regularly and primary tissues degenerate continuously. This new secondary xylem also degenerates the old secondary xylem. Waste materials are formed in the stem such as lignin, suberin, tannin, resin, gums etc. due to degeneration of the cells. All these waste materials are filled in the lumen (cavity) of tracheids and vessels of secondary xylem. Because of this, wood in the central region of the stem becomes dark coloured (Black brown). It is called heart wood or duramen. The peripheral or outer wood which looks light in colour is known as Sapwood or Alburnum. As a result of growing of secondary xylem, the diameter of heart wood increases.
Physiologically active wood is sapwood. The main function of sapwood is water conduction. Heart wood provides maximum mechanical strength to stem.
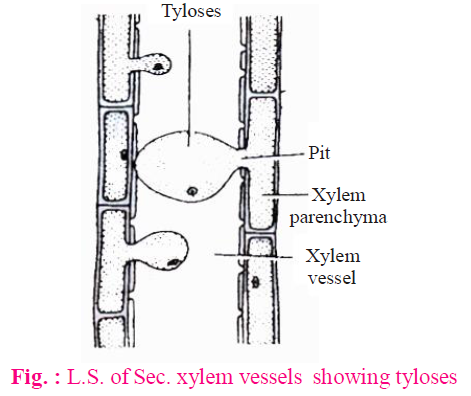
- Conduction of water is not carried by heart wood because :
- Cavities of tracheids and vessels are progressively filled by waste materials.
- The bladder like ingrowth of parenchyma cells enter the lumen of vessels (mainly) & tracheids through pits in their wall. Such bladder like ingrowth are called tyloses. Tyloses blocks the conduction of water.
- In gymnosperms, tylosoids are formed in place of tyloses.

- On the basis of amount of parenchyma, wood is classified into two groups :
- Manoxylic wood : Such type of wood contains more living parenchyma. It is soft and loose wood. E.g., Cycas and other gymnosperm.
- Pycnoxylic wood : Such wood contains less amount of living parenchyma. It is hard wood. Such type of wood are found in most of the plants. E.g., Pinus (Chir), Mango, Acacia (Babool), Tectona (Sagwan), Dalbergia (Shisham).
- On the basis of presence or absence of vessels, wood is classified in two categories-
- Non-porous soft wood : Vessels are absent in such type of wood. E.g., Gymnosperm
- Porous wood : Vessels are present in such type of wood.
- On the basis of arrangement of vessels, porous wood is divided into two groups-
- Ring porous wood : Vessels are arranged in the form of a ring in this type of wood. Such wood conducts water more efficiently.
E.g., Dalbergia. In temperate region.
- Diffused porous wood : Asystematical distribution of vessels is found in this type of wood.
E.g., Azadirachta. In tropical region.
IMPORTANT FACTS
- If the heart wood is destroyed in any stem, then there will be no effect on plants (any vital function is not affected), but if the sap wood is destroyed, then the plant will die because conduction of water will be blocked.
- Heart wood provides stiffness to the stem. The waste materials of heart wood are antiseptic in nature. Heart wood is resistant to bacteria and fungus. Heart wood has a power of repelling insects. So, it is resistant to termites and in the rainy season it does not imbibe water. Thus, it is the best quality of wood.
- Study of wood is known as xylotomy.
- If a wood is exposed freely in air then decomposition of sap wood takes place rapidly.
- Position of youngest layer of secondary phloem is just outside the vascular cambium.
- Position of oldest layer of secondary phloem is just inside the primary phloem.
- Position of youngest layer of secondary xylem is just inside the vascular cambium.
- Position of oldest layer of secondary xylem is just outside the primary xylem.
- If xylem is blocked then shoot will die first.
NOTES
Non-porous wood also called as homoxylous wood and porous wood also called as heteroxylous wood.

(b) Activity of ray initial
- Periclinal divisions occur continuously in ray initials of vascular cambium. Some of the cells are formed towards inside and some of the cells are formed towards outside through these divisions. All these cells are made up of parenchyma. Radial lines of cells of parenchyma are formed in the stem. These are called vascular rays or secondary medullary rays.
- Medullary rays are primary and vascular rays are secondary. Both of them conduct water and food in radial direction. Order of development of vascular rays is in both centripetal and centrifugal manner.
(3) Formation of annual rings
- Annual rings are formed due to unequal activity of the vascular cambium.
- The activity of cambium does not remain same, it is changeable in the whole year. Activity of the vascular cambium is affected by physiological and environmental factors.
- In winter or autumn season, the activity of the cambium is less and the secondary xylem or wood formed is not extensive through the vascular cambium. Cells formed during this period are small thick, walled and have narrow lumens. This is called autumn wood or late wood.
- The vascular cambium is highly active in spring or summer season and secondary xylem formed during this period is extensive and cells of secondary xylem are larger, thin walled and have wider lumen. This wood is known as spring wood or early wood.
- The spring wood is lighter in colour and exhibits low density whereas the autumn (or winter) wood is darker and has higher density.
IMPORTANT FACTS
- The autumn and spring wood are formed in the form of rings. The ring of any type of wood is called growth ring. Thus, two growth rings are formed in one year.
- A ring of autumn wood and a ring of spring wood are collectively known as annual ring. Thus, an annual ring consists of two growth rings. The number of annual rings, formed in a tree give the idea of the age of the tree. The study of determination of age of the plant by this technique is called dendrochronology.
- The annual rings are counted from the base of the stem because basal part has maximum annual rings and upper part has less. Therefore, counting from the basal region can give the correct idea.
- More distinct annual rings are formed in that regions where climatic variations are sharp.
- More distinct annual rings are formed in temperate plants.
- Distinct annual rings are not formed in tropical plants. Distinct annual rings are not formed in India except Himalayan regions.
- Least distinct annual rings are formed in seashore regions because the climate remains the same throughout the year.
- More clear annual rings are formed in deciduous plants as compared to evergreen plants (in temperate region).
- In deserts, annual rings are less distinct.
- Annual rings are bands of secondary xylem and xylem rays.
- Sometimes drought conditions prevail during the middle of a growing season resulting in formation of more than one annual rings. These are called pseudo annual rings.
(B) SECONDARY GROWTH IN EXTRA STELAR REGION
- Secondary growth takes place in extra stelar region due to the activity of cork cambium.
- Cork cambium is also known as phellogen or extrastelar cambium (The cells of the cork cambium are rectangular).
- Cork cambium arises from the hypodermis or from the outer layer of cortex because they become meristematic.
Cork cambium is also formed in the form of a single layered ring. It forms secondary tissue in cortical region.
- Cork cambium also divides periclinally. It forms some cells towards the outside (epidermis) and some cells towards the inside (cortex). Those cells formed towards outside, their middle lamella is suberized. Due to this, these cells become dead. These cells are known as cork or phellem. Those cells formed towards the inside are differentiated into parenchyma and may contain chloroplasts. These are called secondary cortex or phelloderm.
NOTES
- Phellogen, phellem and phelloderm are collectively known as periderm. It means the entire secondary tissue in the cortex is known as periderm.
- Cork is formed in high quantity and secondary cortex is in less quantity because activity of cork cambium is more towards outside. The highest activity of cork cambium is in winter (autumn) season.
- Ring of cork cambium remains living only for one year. Each year, a new cambium is formed below the previous cambium. This new cambium is derived from the secondary cortex or phelloderm.
- All the tissues which occur outside the cork cambium are collectively termed as rhytidome. Rhytidome includes cork and tissues which become dead due to the pressure of cork.
- Most of the cells of phellem are dead. But at some places, living cells are also found. Suberin is not deposited in these places.

These places are known as lenticels. Lenticels appear on the outer surface of the plant either in small points or in the form of areas of protuberance.
Lenticels are made up of scattered group of living cells. Usually they are formed below the stomata. These cells are known as complementary cells/complementary tissue.
- Function of lenticels are
- Exchange of gases between plant and atmosphere. Rows (vertical or longitudinal) of lenticels may occur opposite the medullary rays, facilitating the free exchange of gases.
- Transpiration that takes place through the lenticels, is known as lenticular transpiration.
- Adventitious roots on cutting originates from the living cells of lenticels in vegetative reproduction.
NOTES
- Lenticels are found in most of the woody trees but absent in woody climbers.
- Lenticels are mainly found on wood stems and they are never found on leaves. Lenticels are present all over the plant body. They are also present on fruits.
- If lenticels are blocked then root will die first due to lack of food.
- Bark – There are two views about the bark.
Old view : All the tissues situated outside the cork cambium is called bark. According to old view, bark includes only dead tissues.
Modern view : All the tissues situated outside the vascular cambium is called bark. According to modern view, bark includes both living and dead tissues. Bark has two parts- outer layer of bark is called rhytidome and inner layer of bark is secondary phloem.
NOTES
- If complete bark of tree is removed then plant becomes dead due to excessive water loss.
- If bark is removed in the form of ring at the base of main stem then root will die first due to lack of food.
SECONDARY GROWTH IN DICOT ROOT
- First of all, conjunctive tissue becomes meristematic during secondary growth in a dicotyledon root and form separate curved strips of vascular cambium below phloem bundles. Then after, the cells of pericycle lying opposite to protoxylem also becomes meristematic to form additional strips of cambium. In this way, a complete ring of vascular cambium is formed.
- The portion of vascular cambium formed by pericycle is less.
- The main portion of vascular cambium is formed by conjunctive tissue.
- The shape of a ring of vascular cambium is wavy in the beginning, but later on it becomes circular due to the pressure of secondary xylem.
- The portion of vascular cambium formed by conjunctive tissue becomes meristematic first and form the secondary xylem towards the centre. Ultimately, the ring becomes circular by the pressure of secondary xylem (pushing outwards).

- The activity of vascular cambium of root is the same as the activity of vascular cambium of stem. Secondary xylem is formed towards the inner side and secondary phloem is formed towards the outer side by vascular cambium. The portion of vascular cambium which is formed by pericycle is responsible for the formation of pith rays. These are made up of parenchyma. These pith rays are known as primary medullary rays (multiseriate). A few medullary or pith rays are also formed from remaining vascular cambium. These are called secondary medullary rays (uniseriate). Thus, two types of medullary rays are found in the secondary structure of roots.
NOTES
The presence of two types of medullary rays is basic characteristic feature of roots. Only secondary medullary rays are found in stem after the secondary growth. Both of them conduct water and food in radial direction.
- Cork cambium is developed from the pericycle in roots. Cork is formed towards the outside and secondary cortex is formed towards the inner side by the cork cambium. Lenticels are also found in roots but less in number as compared to stem. Cortex completely degenerate in roots after the secondary growth of one or two years. This falls down due to the pressure of cork, whereas in stem, it degenerates after the long duration.
- Secondary growth is essential in roots to provide strength to the growing aerial parts of the plants and fulfill the requirement of water and minerals.
- Annual rings are not formed in roots because these are not affected by the changes of environment.
- Secondary growth is not found in monocot roots.
FUNCTIONS OF SECONDARY MERISTEM
- Healing of wounds : When any plant part gets injured, wound is formed there. Boundary of the wound is raised outside and composed of similar type of living cells (parenchyma) called callus. Living cells of wound are responsible to form a cambium. This is called wound cambium. It is also called inducible cambium This newly formed cambium forms cork towards the outside. This cork covers the wound entirely. Wound cambium is a secondary lateral meristem.
- Abscission : Falling of any plant organ is called abscission.

Abscission takes place due to formation of abscission layer at the base of plant organ and it is composed of parenchyma. ABA induces the formation of abscission layer.
Middle lamella is dissolved in abscission layer during abscission and primary walls also dissolve partially or completely. Sign of leaf fall on stem is called leaf scar. It is a type of wound.The living cells of leaf scars are responsible to form cork cambium, which produces cork. Cork covers the wound. At the site of abscission, protective layer is found which is suberized.
Post a Comment